Waldlaubsänger (Phylloscopus sibilatrix)
JAHRESRHYTHMUS
Datengrundlage: Beobachtungen der Mitglieder des Arbeitskreises Vogelschutzwarte Hamburg seit 1949, Meldungen im online-Portal ornitho.de seit 2011 und Nestkarten der ehemaligen Beringer W. Harms (1), P. Ruthke (†, 21) und C. Stobbe (†, 2). Statistische Berechnungen erfolgten mit dem „R“-Programm, Version 2.15.2 (2012).
Heimzug
Waldlaubsänger sind nachtaktive Langstreckenzieher; der Wegzug in S – SSE-Richtung beginnt Juli/August, dauert bis Anfang September und führt vornehmlich über Italien nach Afrika, wobei offenbar auch größere Anzahlen die Hochalpen überqueren, (Ringfernfunde: 2 x Burkina Faso, 1 x Süditalien, 1 x Niger/ in England beringte: 8 x Italien, 1 x Algerien). Sie gehören zu den wenigen paläarktischen Arten, die außer in der Feuchtsavanne auch im äquatorialen Regenwald von Oktober/November bis Februar/März überwintern (Bairlein et al. 2014). Der Heimzug beginnt nach einer Vollmauser im März und führt überwiegend über Italien wieder nach Mitteleuropa – nur wenige weiter westliche Funde deuten auf einen eventuellen Schleifenzug hin (Zink 1973). Die Ankunft im Berichtsgebiet erfolgt Ende April/Anfang Mai, im letzten Jahrzehnt in der zweiten Aprilhälfte, ausnahmsweise in der ersten Monatshälfte. Die bisher früheste Beobachtung erfolgte am 5.4.1990, Wohldorf (H. Hargens). Die anderen einheimischen Laubsänger kehren früher zurück. Für den Heimzug benötigt die Art ca. 30 Tage, für den Herbstzug dagegen 62 Tage; der Heimzug wird also doppelt so schnell durchgeführt (Stresemann 1955).
Angaben zur Ankunft aus der älteren Literatur:
In Schleswig-Holstein nach Rohweder (1875) „Anfang Mai“; nach Beckmann (1964) „Ende April“.
In Niedersachsen „erscheinen die ersten Vögel zwischen Mitte April und Mitte Mai, im Mittel in der zweiten Aprilhälfte“ (Zang, H. in Zang et al. 2005).
Bremen: Median/1990-2001= 26.4. (21.-29.04., Seitz et al. 2004).
Stade: Median/2005-2018= 22.04. (10.04.-04.05.; Orn. Jahresberichte).
Raum Hamburg:
Zeitraum Mittelwert/Median Spanne Autor
1876, 1878,1881 27.04. 21.04.-14.05. Böckmann in Krohn, 1924
„aus 18 Daten“ 28.04. Dietrich, 1928
1926-1936 04.05./02.05. 19.04.-27.05. Hennings, 1937
1949-1957 25.04./20.04. 13.04.-29.04. Bruns, 1961
Alle Arbeitskreisdaten aus neuerer Zeit ergeben folgende Ankunftstermine:
Erstbeobachtung Ankunft 20. Ind.
Zeitraum Median/Spanne Median/Spanne
1) 1966-1976;
1984-1995 22.04. (05.04.-29.04.) 03.05.(22.04.-09.05.) (n=23)
2) 1996-2018 15.04. (10.04.-25.04.) 26.04.(19.04.-08.05.) (n=23)
3) 1966-1976;
1984-2018 19.04. (05.04.-29.04). 29.04. (19.04.-09.05.) (n=46)
Verfrühungen (n=46):
Erstbeobachtung: -0,15212** Tage/Jahr p<0,01) = -7,0 Tage
20. Individuum: -0,19103*** Tage/Jahr p<0,001) = -8,8 Tage
Letztbeobachtungen:
1) 21.07. (14.06.-19.09.)
2) 23.07. (13.06.-11.09.)
3) 23.07. (13.06.-11.09.)
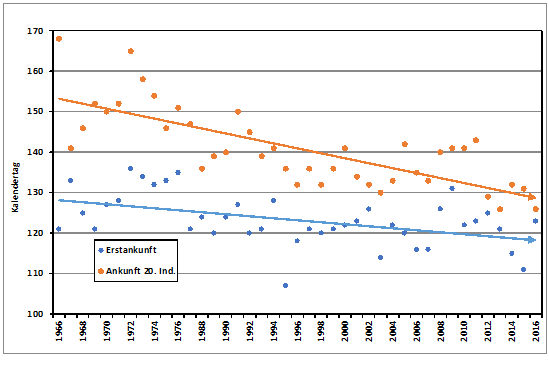
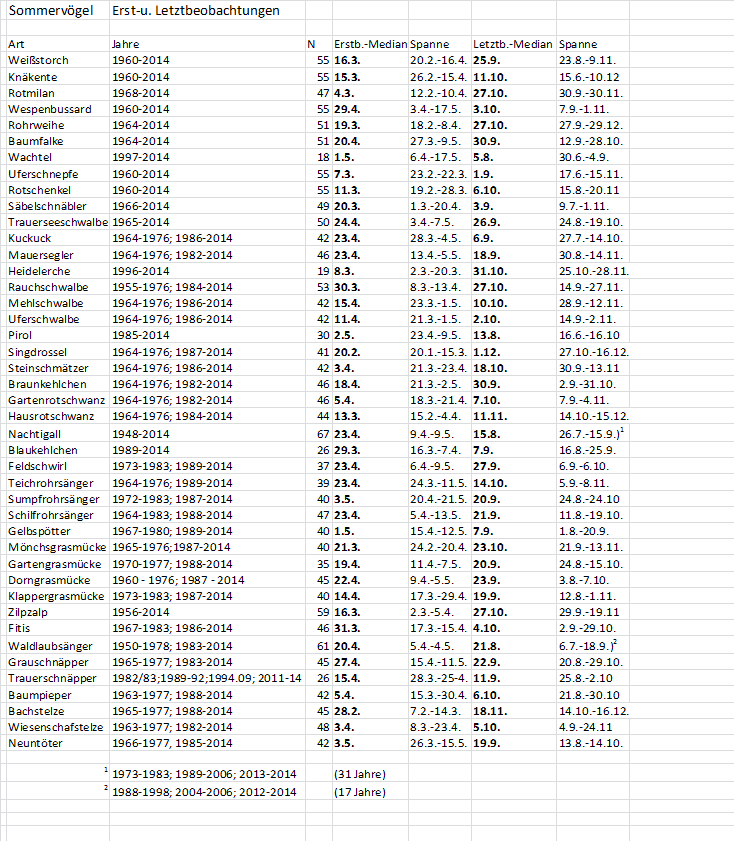
Zur Ankunft in den letzten Jahrzehnten stehen ausreichende Beobachtungsmeldungen zur Verfügung aus den Jahren 1966-1976 und 1984-2018. Median der Erstbeobachtungen aus diesen 46 Jahren ist der 19.04. (05.04.-29.04.). Die Ankunft der gesamten Brutpopulation kann sich aber noch über mindestens 4 Wochen hinziehen (Glutz von Blotzheim & Bauer 1991). Das 20. Individuum kehrte im Mittel am 29.04. (19.04.-09.05.) in das Hamburger Berichtsgebiet zurück. Die Ankunft der ersten Waldlaubsänger verfrühte sich im genannten Zeitraum signifikant um 6,4 Tage, die des 20. Individuums hoch signifikant um 8,8 Tage (Abb. 1). Zwischen den zwei Ankunftsreihen Erstbeobachtung und 20. Individuum besteht eine hochsignifikante Korrelation. In Berlin ist der Erstbeobachtungsmedian der 16.04. (04.04.-29.04), die Verfrühung betrug (1966-1976,1984-2018)1 lediglich 6,8 Tage (schwach signifikant). Die Ankunftsreihen beider Städte sind hoch signifikant miteinander
korreliert. Im sächsischen Voigtland wurde eine Verfrühung der Erstankunft um 14,1 Tage (1967-2016) festgestellt (Friedel & Ernst, 2017). In Hessen hat sich (2007) der Brutbeginn um 8 Tage verfrüht (Hillig 2009); eine entsprechende Ankunftsverfrühung ist zu vermuten.
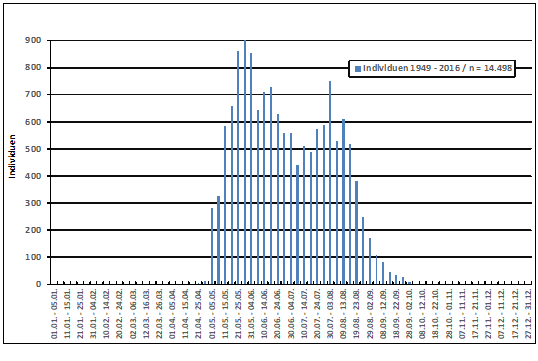
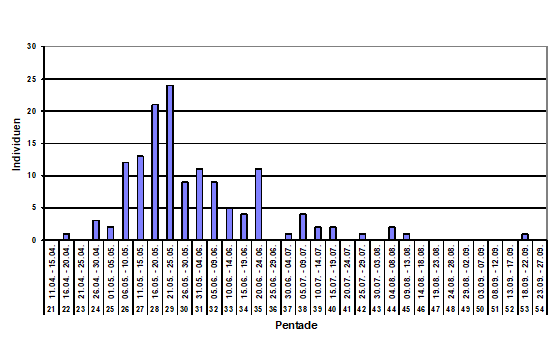
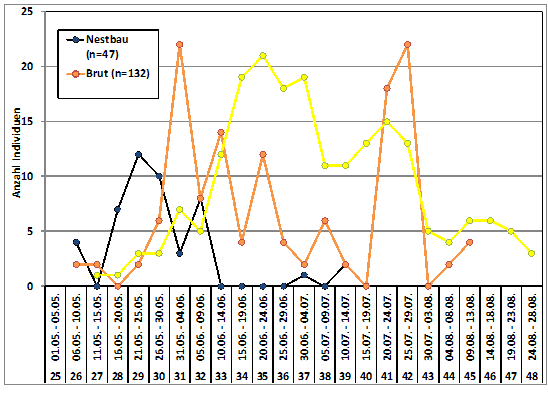
Zum Heimzug gibt es keine Beobachtungen; nach Erstfängen in der Beringungsstation „Die Reit“ wurden 1984-1988 zwischen dem 22.4. und 4.6. 10 Individuen gefangen (Dinse 1991). Höchste Individuenzahlen und Maxima der Gesangsaktivität werden aus der 25./26. Pentade (01.05.-10.05.) gemeldet; die Art wird überwiegend akustisch registriert (Abb. 2). Ein zweiter kleinerer Gipfel ist möglicherweise mit nordischen Durchzüglern zu erklären.
1 ohne 2004 und 2009; es gab keine Datenangabe in Berlin
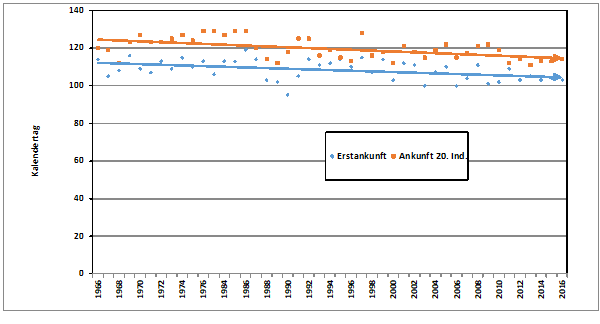
Abb. 1: Waldlaubsänger – Erstbeobachtung und Ankunft 20. Individuum im Raum Hamburg
(1966-1976, 1984-201ß)
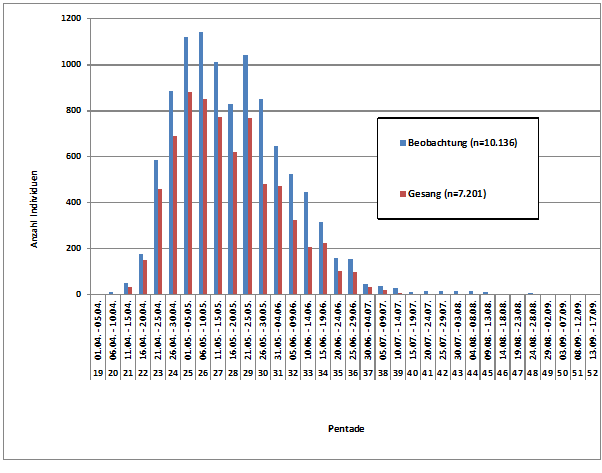
Abb. 2: Waldlaubsänger – Jahreszeitliches Auftreten und Gesangsaktivität
im Raum Hamburg (1948 – 2018)
Brut
Unmittelbar nach der Ankunft werden die Reviere besetzt; nach Mühlenberg (1964) treffen die Weibchen ca. 3 Tage später am Brutplatz ein. Eine extrem schnelle Revierbesetzung wurde 1985 in Schottland dokumentiert (Morton 1986): Ein am 08.05. auf der Isle of Man beringtes Männchen wurde am 09.05. in Schottland (200 Km nördlich) singend im Revier angetroffen. Abends wurde es schon zusammen mit einem Weibchen beim Nestbau beobachtet. Kurz darauf konnte es gefangen und der Ring abgelesen werden. Das Nest war am 13.05. fertig gebaut.
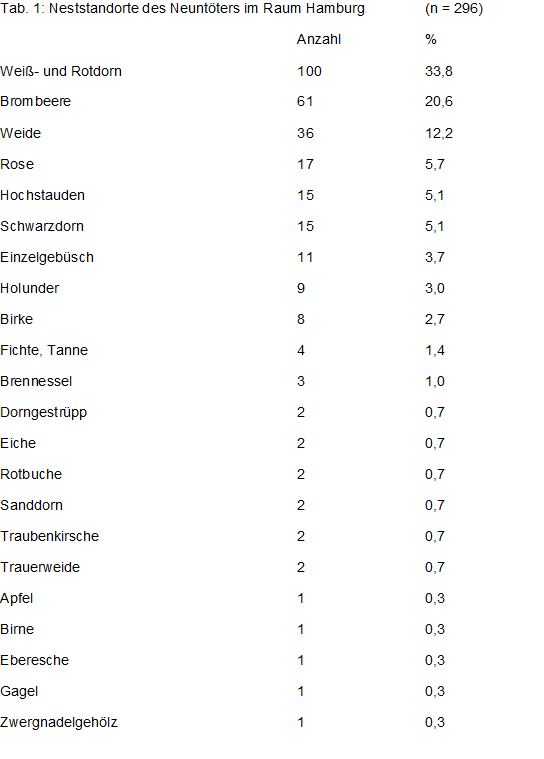
Waldlaubsänger bevorzugen Nadel-Laub-Mischbestände (Buche, Eiche, Fichte bzw. Kiefer mit Laubhölzern gemischt), Deckungsgrad 70-80 %, vor allem Baumholz, seltener Stangen- oder Altholz. Im Berichtsgebiet wird als häufigste Habitatangabe ebenfalls Laubmischwald gemeldet; immerhin 14,5 % stammen aus waldartigen städtischen Grünanlagen, s. Tab 1. Wichtig ist eine schwach entwickelte Strauchschicht bzw. Baumäste müssen bis auf 2,5 m herabreichen, damit der Nestanflug von dort möglich ist. Der Deckungsgrad muss aber kleiner als 40 % sein um den Singflug des Männchens nicht zu behindern. Günstig ist ein beständiges Kleinklima. Das Nest wird gerne in Grasbulten angelegt, oft aber auch in altem Laub. Bevorzugt werden Südhanglagen (z.B der SE-Hang am Schüberg/OD) oder windgeschützte Waldränder (Quelle & Tiedemann 1972, Tiedemann 1970, 1971). Revierkartierer im Berichtsgebiet (Berg, Mulsow) stellten häufig fest, dass im Laubmischwald Stellen mit einzelnen Nadelhölzern bevorzugt werden und umgekehrt.
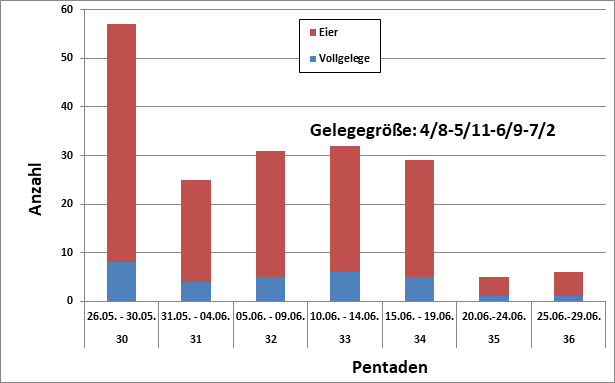
Revierverhalten, Balz und Nestbau wurden im Berichtsgebiet ab 23.04. gemeldet, die letzte Beobachtung vom 20.07.; Median 107 = 28.05. Daten zum Legebeginn liegen vor vom 13.05. bis 05.06.; Median 22 =22.05. In Niedersachsen wurde im Mittel der 27.05. (n=8) gefunden (Keßler in Zang 2005); frühester Termin im Rheinland war der 04.05. (Glutz von Blotzheim & Bauer 1991). Die mittlere Eigröße betrug n. Dietrich im Berichtsgebiet 15,3 x 12,2 mm; in Mitteleuropa nach Makatsch in Glutz von Blotzheim & Bauer 1991: 16,3 x 12,8 mm.
Gelegegrößen im Raum Hamburg (25 Nestkarten):
2 3 4 5 6 7
1 3 0 10 10 1; im Mittel also 5,1 Eier/Nest.
Dietrich 1928 gibt für Hamburg 6-7 Eier an. Für Mitteleuropa werden 4-8, meist 6 Eier angegeben (Glutz von Blotzheim & Bauer 1991). In NRW fand Mildenberger (1940) für 132 Maigelege ein Mittel von 6,15, für 78 Juni-Vollgelege 5,46 Eier. Niedersachsen meldet 6,o2 (n=63) n. Smith in Zang 2005; Brandenburg 6,1 (n=16) n. Rutschke 1983. In der Regel findet nur eine Jahresbrut statt, bei Gelege- oder Brutverlusten werden aber regelmäßig Ersatzgelege getätigt.
Brütende Waldlaubsänger wurden meistens aus der zweiten Maihälfte gemeldet (24x), seltener aus der ersten Junihälfte (5x). Die früheste Meldung „Eier bebrütet“ erfolgte vom 04.05.1969, Giesensand (H. Vidal), die späteste vom 14.06.2009, Wohldorfer Wald (E. Paulsen). Die Termine für schlüpfende Nestlinge liegen zwischen dem 01.06. und 12.07.; Median 29 = 29.06.
Gemeldete Anzahl der Nestlinge:
0 1 2 3 4 5 6 7
1 0 1 4 0 9 9 1; Mittel: 4,8 Jungvögel/Nest.
Nach Beringungslisten aus dem Hamburger Raum betrug die durchschnittliche Brutgröße 5,4 Nestlinge (n=36); beringt werden Nestlinge im Alter von 6-12 Tagen (Beringer: Alpheis, H., Bosselmann, F., Gruner, D., Hinze, P.,Ruthke, P., Schneider, A., Steppan, W.).
In Niedersachsen wurden 5,8 Jungvögel/Nest ermittelt (Keßler in Zang 2005); in Brandenburg 5,4 (Rutschke 1983). Für Mitteleuropa werden 4,4 Jungvögel/Nest genannt (Glutz von Blotzheim & Bauer 1991). Eiablage, Brut und Nestlingszeit beanspruchen ca. 33 Tage. Nachgelege gibt es bis Anfang Juli. Echte Zweitbruten sind sehr selten (Stresemann 1955).
Flügge Jungvögel wurden im Raum Hamburg vom 25.05. bis 27.07. beobachtet (n=23); Median: 23.06. Nach den vorliegenden Nestkarten war der Bruterfolg mit 93,7 % sehr hoch, wobei es sich überwiegend um Beringungsdaten handelt; große Verluste erfolgen ja meist erst danach unter den flüggen Jungvögeln. Es gab einen Totalverlust durch Prädatoren, zwei tote Jungvögel und ein taubes Ei. Für Mitteleuropa wird eine Ausfliegerate zwischen 59 und 78,5 % angegeben (Glutz von Blotzheim & Bauer 1991). Neben schlechter Witterung sind vor allem Wildschweine, Marder, Igel, Mäuse, Eichhörnchen u.a. für z.T. hohe Verluste verantwortlich. Lokal kann es geringe Verluste durch Kuckucksparasitierung geben; nach Soldat (1981) wurden 1978-1980 29 % von 62 Nestern vom Kuckuck belegt, aber nur 4 Kuckuckseier wurden ausgebrütet. Für Eiablage, Brut und Nestlingzeit rechnet Stresemann (1955) mit 33 Tagen. Nachgelege können bis Anfang Juli erfolgen. Der Aufenthalt im Brutgebiet beträgt ca. 100 Tage.

Wegzug
Waldlaubsänger ziehen ca. 2 Wochen früher weg als der Fitis, der eine Vollmauser durchführt, während Waldlaubsänger nur das Kleingefieder mausern. Nach Gwinner (1969) beginnt die Zugunruhe durchschnittlich im Alter von 60; beim Fitis von 65 und Zilpzalp von 125 Tagen. Die Zerstreuung der Jungvögel setzt Ende Juni/Anfang Juli ein. Die Fangzahlen in der Beringungsstation „Die Reit“ sind so gering, dass sich daraus keine gesicherten Aussagen ableiten lassen. 1974-1983 wurden insgesamt 6 Individuen nach dem 30.07. gefangen (Berthold et al 1991); seit 1995 gingen 9 von 17 vorher ins Netz. Das wäre ein Hinweis für eine mögliche Verfrühung beim Wegzug. Nach Fängen in Serrahn (Mecklenburg-Vorpommern, geographische Breite etwa wie Hamburg) ziehen vermauserte Jungvögel ab 20.07. weg, Altvögel nach dem 10.08. bis Anfang September (Starke & Weber in Klafs & Stübs 1977). Während Fitis und Zilpzalp aus Norddeutschland als SW-Zieher gelten, wandern Waldlaubsänger vorwiegend in SSE-Richtung ab. Nach Stresemann (1955) gelangen Vögel aus Europa alle in einen Zugtrichter, der vom Pyrenäenostrand bis Westgriechenland reicht. Über Italien und das zentrale Mittelmeer gelangen sie nach Nordafrika und die Art ist “ Anfang August einer der häufigsten Vögel in Tunesien“. Die Überwinterungsgebiete liegen in einem Streifen zwischen Äquator und 10 ° Nord von Westafrika (Goldküste) bis zum Ostrand des Kongobeckens. Dort erfolgt eine Vollmauser von Ende Dezember bis März, worauf der Heimzug beginnt.
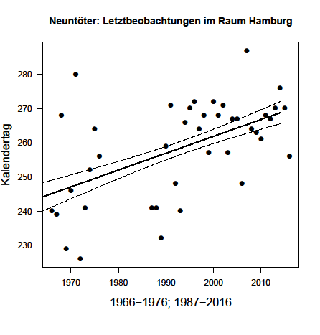
Das Wegzugmaximum liegt im August, aber Jungvögel aus späten Ersatz- oder Zweitbruten können noch im September beobachtet werden, Einzelvögel ausnahmsweise im Oktober/November. Im Hamburger Raum fällt der Median der Letztbeobachtungen im genannten Zeitraum auf den 23.07. (13.06.-06.11.). Eine Verspätung von 15,6 Tagen über 44 Jahre ist statistisch nicht signifikant (Abb. 3). Median der Letztbeobachtungen in Berlin ist der 30.8. (23.07.-17.09.); die Verspätung beträgt 5,7 Tage.
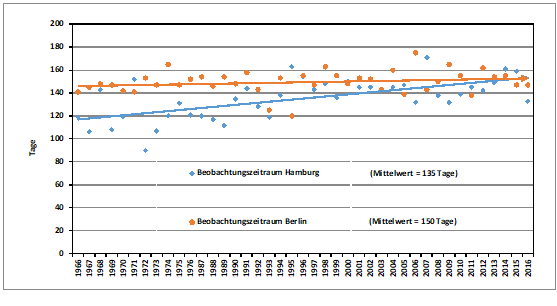
Der Beobachtungszeitraum in Hamburg von durchschnittlich 98 +/- 28 Tagen (54-148) hat sich in 44 Jahren (schwach signifikant) um 21,9 Tage verlängert. In Berlin ist der mittlere Beobachtungszeitraum von 136 +/-14 Tagen um 12,5 Tage größer geworden (Abb. 4).
Abb. 3: Waldlaubsänger – Letztbeobachtungen im Raum Hamburg
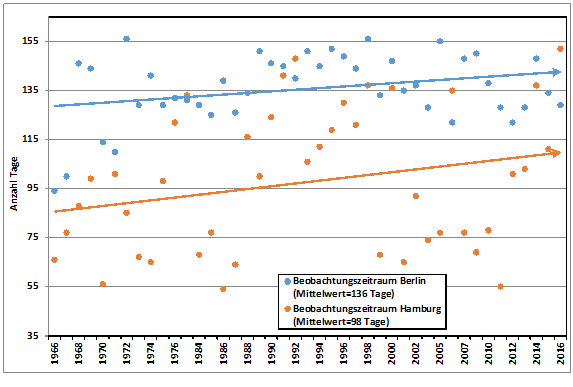
Abb. 4: Waldlaubsänger – Beobachtungszeiträume in Berlin und Hamburg
Literatur:
Bairlein, F. & J. Dierschke, V. Dierschke, V. Salewski, O. Geiter, K. Hüppop, U. Köppen, W. Fiedler (2014): Atlas des Vogelzugs. Aula-Verlag Wiebelsheim. 567
Bertholdt, P., Fliege, G., Heine, G., Querner, U. & R. Schlenker (1991): Wegzug, Rastverhalten, Biometrie und Mauser von Kleinvögeln in Mitteleuropa.- Vogelwarte 36 (Sonderheft): 1-221.
Dinse, V. (1991): Über den Heimzug von Kleinvögeln in Hamburg. Eine Auswertung im Rahmen des Mettnau-Reit-Illmitz-Programms.-Hamburger avifaun. Beitr. 23: 1-125.
Friedel, W. & S. Ernst (2016): 50 Jahre Beobachtung, Dokumentation und Auswertung der Erstankunftstermine heimkehrender Zugvögel im Vogtland – eine Analyse zwischen 1967 und 2016. Mitt. Ver. Sächs. Ornithol., im Druck.
Glutz von Blotzheim, U. N. & K. M. Bauer (1991): Handbuch der Vögel Mitteleuropas. Bd. 12, Teil 3. Aula-verlag, Wiesbaden.
Gwinner, E. (1969): Untersuchungen zur Jahresperiodik von Laubsängern. J. Orn. 110: 1-21.
Hillig, F. (2009): Der Bestandsrückgang des Waldlaubsängers. Falke 56: 60-63.
Keßler, A. (1963): Beobachtungen an einer Waldlaubsängerpopulation (Phylloscopus sibilatrix) im Bloher „Wold“ unter besonderer Berücksichtigung ökologischer und ethologischer Verhältnisse. Pädagog. Hochschule Oldenburg.
In: Zang, H., H. Heckenroth & P. Südbeck (2005): Die Vögel Niedersachsens, Drosseln, Grasmücken, Fliegenschnäpper. Naturschutz Landschaftspfl. Niedersachs. B, H 2.9.
Makatsch, W. (1976): Die Eier der Vögel Europas. Neumann Verlag.
Mildenberger (1940): Beobachtungen über Fitis-, Weiden- und Waldlaubsänger im Rheinland. J. Orn. 88: 537-549.
Morton, R. (1986): Rapid territory establishment by a Wood Warbler. Ringing & Migration 7: 56.
Mühlenberg, M. (1964): Brutzeitbeobachtungen am Waldlaubsänger. Vogelwelt 85: 174-182.
Otto, W. (Artbearbeiter) in Rutschke, E. (1983): Die Vogelwelt Brandenburgs, Jena 1983
Quelle, M. & G. Tiedemann (1972): Strukturanalyse von Waldlaubsängerrevieren im Raum Bielefeld. Abh. Landesmus. Naturkde Münster Westfalen 34: 95-102.
Seitz. J., K. Dallmann und T. Kuppel (2004): Die Vögel Bremens und der angrenzenden Flußniederungen. 416 S.
Smith, A. H. V. (1958): Excess of male Wood Warblers. Bird Study 5: 113.
In: Zang, H., H. Heckenroth & P. Südbeck (2005): Die Vögel Niedersachsens, Drosseln, Grasmücken, Fliegenschnäpper. Naturschutz Landschaftspfl. Niedersachs. B, H 2.9.
Starke, W. & H. Weber in Klafs, G. & J. Stübs (1977): Die Vogelwelt Mecklenburgs. Fischer Jena.
Stresemann, E. (1955): Die Wanderungen des Waldlaubsängers (Phylloscopus sibilatrix). J. Orn. 96: 153-167.
Tiedemann, G. (1970): Waldlaubsänger. J. Orn. 111: 277.
Tiedemann, G. (1971): Zur Ökologie und Siedlungsdichte des Waldlaubsängers (Phylloscopus sibilatrix). Vogelwelt 92: 8-17.
Zang, H., H. Heckenroth & P. Südbeck (2005): Die Vögel Niedersachsens, Drosseln, Grasmücken, Fliegenschnäpper. Naturschutz Landschaftspfl. Niedersachs. B, H 2.9.
Zink, G. (1973): Der Zug europäischer Singvögel. Vogelwarte Radolfzell Bd1, 1973.
(Hamburg im November 2020)
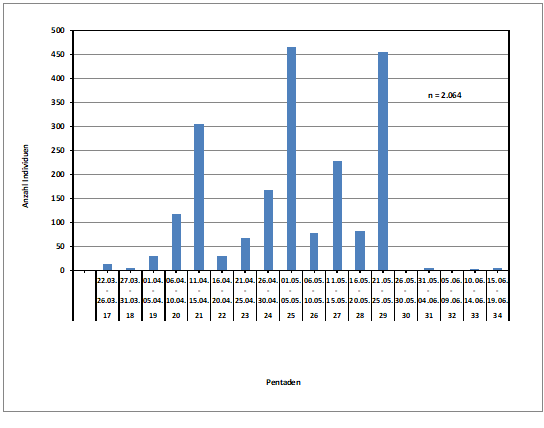
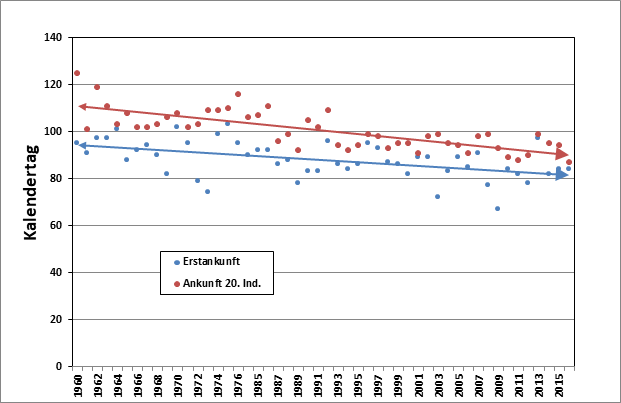
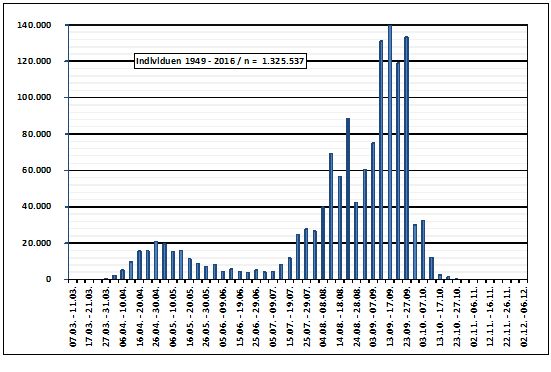
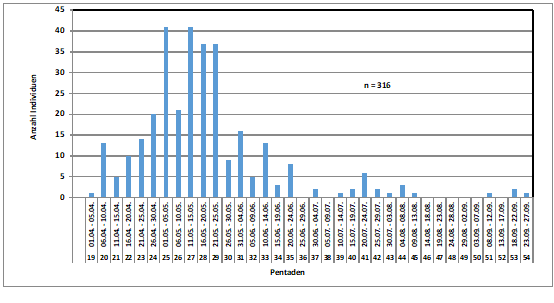
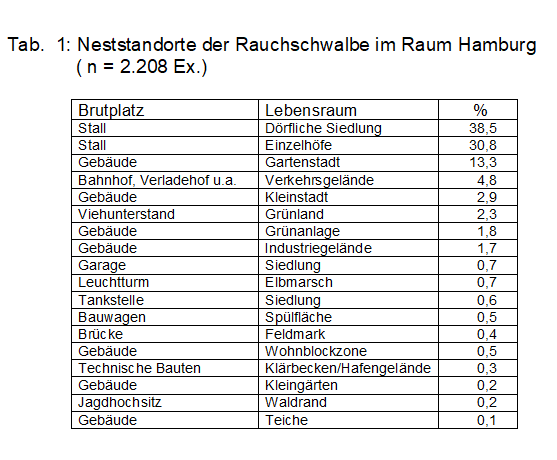
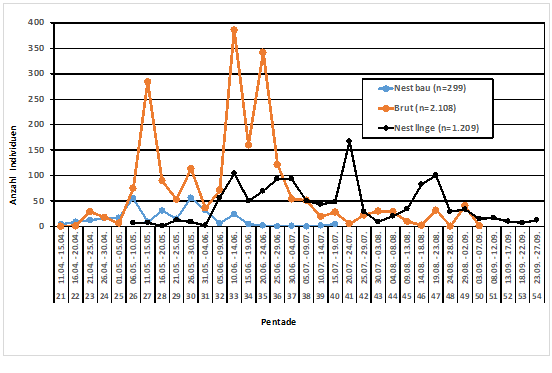 Abb. 5: Rauchschwalbe – Brutaktivitäten im Raum Hamburg
Abb. 5: Rauchschwalbe – Brutaktivitäten im Raum Hamburg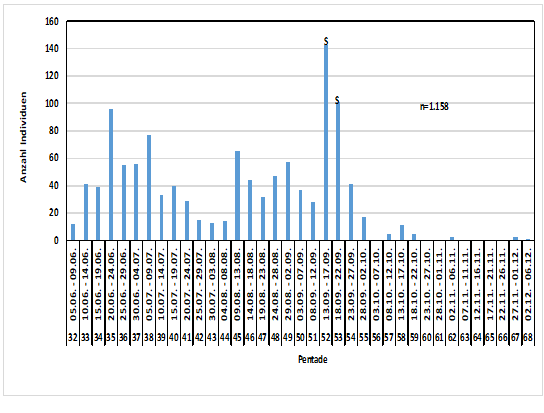
 Abb. 7: Wegzug der Rauchschwalbe im Raum Hamburg (1948-2016)
Abb. 7: Wegzug der Rauchschwalbe im Raum Hamburg (1948-2016)