Datengrundlage: Beobachtungen der Mitglieder des Arbeitskreises Vogelschutzwarte Hamburg seit 1949, Meldungen im online-Portal ornitho.de seit 2011, Nestkarten der ehemaligen Beringer D. Gruner (1), P. Hinze (66) und P. Ruthke (6). Statistische Berechnungen erfolgten mit dem „R“-Programm, Version 2.15.2 (2012).
Heimzug
Neuntöter gehören zu den wenigen Langstreckenziehern unter den Singvögeln, die über das östliche Mittelmeer ihre Winterquartiere in Ost- und Südafrika erreichen; selbst die westlichsten Populationen nehmen diese Route (Ringfunde von Nestlingen, die bei Bad Oldesloe beringt wurden: Italien 1x, Griechenland 1x, Ägypten 1x). Diese Arten (z. B. auch Klappergrasmücke und Sumpfrohrsänger) kehren daher relativ spät in ihre Brutgebiete in Mitteleuropa zurück. Für Neuntöter ist zudem ein Schleifenzug, entgegen dem Uhrzeigersinn, über die arabische Halbinsel nachgewiesen (Bairlein et al. 2014).
Die Ankunft im Berichtsgebiet erfolgt je nach Witterung Ende April/Anfang Mai; gegen Ende Mai ist in der Regel die gesamte Brutpopulation eingetroffen. Die bisher früheste Erstbeobachtung (nach Daten des Arbeitskreises Vogelschutzwarte Hamburg) wurde am 17.04.1995, Mühlensand (U. Rahr) nach einem Mildwinter gemeldet.
Angaben zur Ankunft in der älteren Literatur: In Schleswig – Holstein nach Rohweder (1875) „anwesend von Anfang Mai bis Ende August“. In Niedersachsen kommt die Art in der ersten Maihälfte an (Zang in Zang et al. 1998); in östlichen Landesteilen 05. – 07.05., in westlichen 13. – 15.05. Ankunft in Hamburg (Bruns 1961) für die Jahre 1949 – 1957: 11.05.; Spanne: 14.04. – 23.05.
Aus neuerer Zeit stehen für eine Auswertung ausreichende Beobachtungsmeldungen zur Verfügung für die Jahre 1966 – 1976 und 1987 – 2016. Der Median für die Erstbeobachtungen in diesen 41 Jahren ist der 02.05. (17.04. – 16.05.). Das 20. Individuum wird dagegen im Mittel erst am 20.05. (06.05.-17.06.) im Raum Hamburg gemeldet. Im genannten Zeitraum verfrühte sich die Ankunft der Erstbeobachtungen nach der Regressionsanalyse hoch signifikant (p < 0,001) um 9 Tage, die des 20. Individuums sogar um 20 Tage. Damit ist erwiesen, dass die Verfrühungen nicht etwa nur die ersten Vögel betreffen, sondern deutlich noch stärker das Gros der Population, s. Abb.1. Die Korrelationen zwischen den Ankunftsreihen sind hoch signifikant. Die Erstbeobachtungsdaten zeigen eine signifikante Korrelation (Spearman-Rangkorrelation) mit den April-Mitteltemperaturen von Thüringen, Sachsen und Sachsen-Anhalt (r = 0,50**). Dieses Ergebnis entspricht der Zugroute dieses Südostziehers. Im sächsischen Vogtland wurde eine Verfrühung der Erstankunft um 11,2 Tage (1967 – 2016) festgestellt (Friedel, W. & S. Ernst 2017 im Druck). Die mittlere Erstankunft war dort in 50 Jahren der 5.5. (13.04.-18.05.). In Berlin betrug die Verfrühung lediglich 6,6 Tage (1965-2016), nach Daten der Berliner ornithologischen Arbeitsgemeinschaft); Median der Erstbeobachtung: 03.05. (25.04.-15.05.). Mehrere Autoren betonen die Abhängigkeit der Erstankunft von der mittleren Apriltemperatur (Ernst 2013, Münster 1958, Stresemann 1944, Tauchnitz 1968). Die Hamburger Ankunftsdaten zeigen eine hochsignifikante Korrelation mit den April-Mitteltemperaturen von Sachsen (r=0,50***; Spearman-Korrelation); signifikant sind ebenso die Werte für Thüringen (r=0,50**) und Sachsen-Anhalt (r=0,49**). Dieses Ergebnis passt gut zur Zugroute dieses Südostziehers.
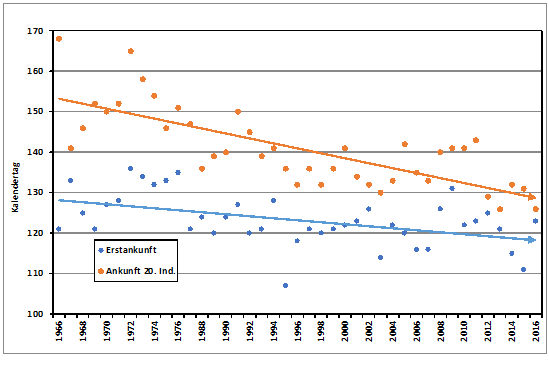
Abb. 1: Neuntöter – Erstbeobachtungen und Ankunft des 20. Individuums im Raum Hamburg / 121 = 1. Mai
Ankunftsdaten für verschiedene Zeiträume:
Erstbeobachtung Zweitbeobachtung Ankunft 20. Indiv.
Zeitraum Median/Spanne Median/Spanne Median/Spanne
1. 1966-1976;
1987-1995 06.05./17.04.-16.05. 20.05./30.04.-24.05. 27.05./16.05.-17.06.
2. 1996-2016 01.05./24.04.-11.05. 03.05./26.04.- 12.05. 13.05./06.05.-21.05.
3. 1966-1976;
1987-2014 02.05./17.04.-16.05. 05.05./26.04.-24.05. 20.05./06.05.-17.06.
Verfrühungen (n=41):
Erstbeobachtung: 0,21*** Tage/Jahr (R²=0,28; p<0,001) = 8,8 Tage
Zweitbeobachtung: 0,26*** Tage/Jahr (R²=0,42; p<0,001) = 10,8 Tage
20. Individuum: 0,49*** Tage/Jahr (R²=0,62; p<0,001) = 20,2 Tage
Der Ankunftsort lag in den letzten 20 Jahren 11mal im Ost-, 8mal im Süd- und 1mal im Nordwestteil des Berichtsgebietes, was bei einem „SO-Zieher“ zu erwarten ist. Vom Frühjahrszug wurden nur wenige Beobachtungen dieses Nachtziehers gemeldet:
1 Ind. 09.05. Escheburger Moorwiesen/RZ, Zug nach O (Nehlsen 1982)
5 Ind. 16.05. Escheburger Moorwiesen/RZ, Zug nach O (Nehlsen 1981)
1 Ind. 18.05. Neuer Hetlinger Koog /PI (Hahn 1997)
1 Ind. 22.05. Ohmoor (Bruster 1996)
1 Ind. 28.05. Süderelbemoore, Zug nach N (Nehlsen 1983)
1 Ind. 29.05. Junkernfeldsee/WL, Zug nach N (Sannow 1994)
In der Regel kommen die Männchen 1 – 2 Tage vor oder zusammen mit den Weibchen an (Jakober & Stauber 1983). Im Raum Hamburg war es 24mal ein Männchen, 5mal beide, 1mal ein Weibchen, 14mal unbekannt (n=44).
Zwei Gebiete wurden so häufig besucht, dass die geschlechtsspezifische Zeitverschiebung bestätigt werden konnte: Am 30.04. wurden zwei Männchen und am 02.05. zwei Paare im Duvenstedter Brook (Hargens, 1989) sowie in Altengamme am 01.05. das Männchen und am 04.05. das Weibchen (Lade, 1993) beobachtet. Nach Untersuchungen an einer buntberingten Population in Süddeutschland kommen spät wegziehende Individuen im Frühjahr auch später an (Jakober & Stauber 1983). Die Brutortstreue soll bei Männchen etwas grösser sein, ist aber insgesamt relativ niedrig (Jakober & Stauber 1989, Simek 2001).
Der Einzug der Brutpopulation ist gegen Ende Mai abgeschlossen; aus der 30. Pentade (26.05.-30.05.) wird die höchste Individuenzahl des Jahres gemeldet (wohl inklusive nordischer Durchzügler); der zweite Gipfel Anfang August wird vermutlich durch die ausgeflogenen Jungvögel bestimmt, s. Abb. 2 und Abb. 6.
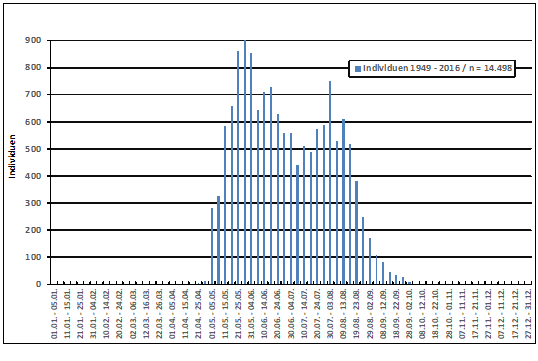
Abb. 2: Neuntöter – Jahreszeitliches Auftreten im Raum Hamburg (1949 – 2016)
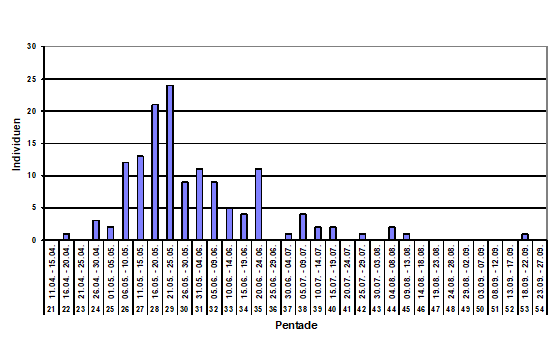
Abb. 3: Neuntöter: Gesangsaktivität (1949-2016; n=138, Median: 29. Pentade)
Brut
Revier: Direkt nach der Ankunft im Brutgebiet beginnen die Neuntöter mit dem Revierverhalten, also Gesang, Revierkämpfe oder Nestbau. Im Berichtsgebiet wurde Gesangsaktivität ab dem 17.04. (Rahr 1995) gemeldet; das Maximum fällt in die 29. Pentade (21.-25.05.)), es endet gegen Anfang August, s. Abb. 3. Ein sehr später „Herbstgesang“ wurde am 21.09. (Grams 1975) registriert.
Nest: Früheste Nestbau – Meldungen gibt es aus der ersten Maidekade: 08.05.2006, Schwarze Kate-Süd/OD (Berg). Das Maximum wird Ende Mai/Anfang Juni erreicht; die letzte Meldung: 04.07.1973, Schlappenmoor/SE (Dietrich), s. Abb. 5. Neuntöter pflegen Beutetiere, die sie nicht gleich verzehren, auf Dornen aufzuspießen. Bevorzugte Neststandorte sind daher dorn- oder stacheltragende Sträucher (im Raum Hamburg ca. 67 % aller Nester) insbesondere Weißdorn, Schlehe und Brombeere, vereinzelt auch Bäume, s. Tab. 1. Besonders beliebt sind Einzelbüsche oder Feldhecken (Knicks), gerne an Böschungen oder Gräben in halboffener Weidelandschaft mit extensiver Nutzung. Der Gehölzdeckungsgrad sollte höchstens 10-15 % betragen (Jakober $ Stauber 1981, Schuler 1982, Schönfeld 1986, Brandl et al. 1986, Mann 1987, Kowalski 1995, Wollscheid 1995, Vizslan & Pingitzer 2000, Vanhinsberg & Evans 2002, Tryjanowsli et al. 2004, Beiche & Luge 2006, Brambilla et al. 2007). Freistehende Hecken oder Einzelbüsche sind effizienter als Jagdreviere am Waldrand. Die Nesthöhe steht im Zusammenhang mit der Vegetationshöhe. Die Auswertung von Nestkarten ergab M61 = 95,4 cm (0 – 320 cm); darunter waren zwei Bodenbruten. Vegetationshöhe: M53 = 311 cm (70 – 1.200 cm). Der Nestbau beansprucht bei günstiger Witterung 4 – 6 Tage, sodass ca. 8 Tage nach der Ankunft, erfolgreiche Verpaarung vorausgesetzt, das erste Ei gelegt wird.
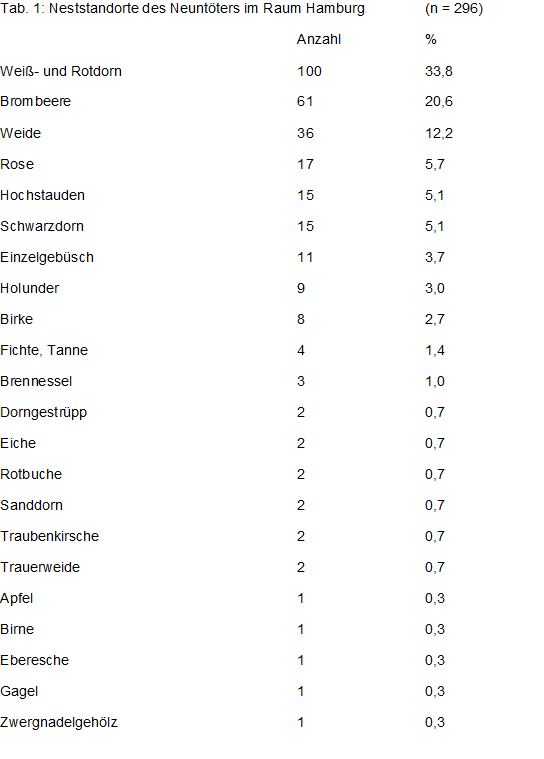
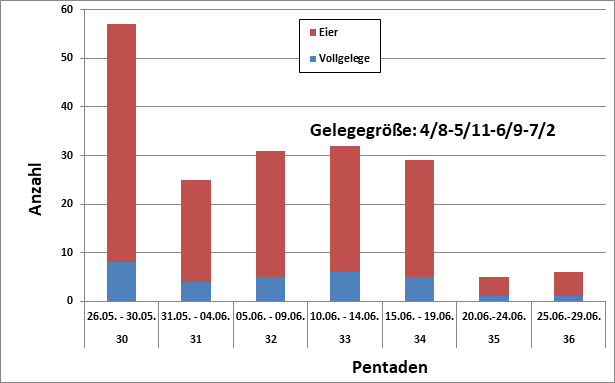
Abb. 4: Neuntöter: Erstbruten – Vollgelege und Gelegegröße (n=30/155, Median: 32. Pentade)
Gelege: Die meisten Vollgelege wurden Ende Mai registriert, s. Abb. 4. In Sachsen-Anhalt lagen die frühesten Termine für Legebeginn zwischen dem 10. Und 20.5. gefunden; ca. 50 % Ende Mai; der rest Anfang bis Mitte Juni (Stein 1972). Die Gelegegröße variiert von 2 – 7, M30 = 5,16 Eier/Nest bei Erstbruten im Berichtsgebiet, bei Ersatzgelegen wurden M8 = 4,6 Eier/Nest gefunden. Dies entspricht weitestgehend den Angaben in der Literatur (Schreurs 1971,Ullrich 1971, Poltz 1975, Luder 1986, Kuzniak 1991, Glutz von Blotzheim 1993). Mittlere Eigröße im Hamburger Raum M170 = 22,08×16,75 mm nach Moebert in Glutz von Blotzheim 1993 (M4 = 22,75×17,25 nach Dietrich 1928). Im Berichtsgebiet war der Legebeginnmedian nach Nestkarten M39 = 07.06. (18.05.-25.06.), zurückgerechnet auf der Basis angegebener Eizahlen bzw. Alter der Nestlinge, in Süddeutschland M24 = 14.05. (06.05.-27.05.) nach Jakober & Stauber (1983). Brütende Neuntöter im Hamburger Raum wurden ab 07.05. gemeldet, Tangstedter Forst (Hasse 1959), mit einem Höhepunkt Anfang Juni (Erstbruten) und einem Nachgelegegipfel in der zweiten Julihälfte, s. Abb. 5. Die Meldungen von späten Nachbruten enden mit dem 28.07., Wakendorfer Moor/SE (Carstens 1979). Die Bebrütung beginnt nach dem vorletzten Ei und dauert 14 – 16 (18) Tage, Männchen und Weibchen brüten. Ei- bzw. Nestverluste können nach den Angaben der Beringer (u. a. Nestkarten von Hinze) bis über 40 % betragen und beruhen vor allem auf Prädation, Witterung und menschliche Einflüsse; ähnliche Ursachen werden in der Literatur bestätigt (Jakober & Stauber 1987, Kuzniak 1991, Knysh 1994, Beiche & Luge 2006). Einzelbüsche werden intensiv von Krähenvögeln durchsucht und Greifvögel erbeuten Altvögel.
Nestlinge: Der berechnete Schlupftag lag für die meisten Jungvögel aus Erstbruten in der 33. Pentade (10.06.-14.06.) mit M28 = 4,75 Jungvögel/Nest nach aus Nestkarten berechneten Daten vom 11.06.-03.07., (2×2, 3×3, 7×4, 7×5, 7×6, 2×7 Jungvögel/Nest). Bei den Nachgelegen (05.07.-05.08.) schlüpften die meisten Nestlinge in der 38. Pentade (05.07.-09.07.).
Die erste Meldung von einem Nest mit nicht flüggen Jungvögeln stammt vom 12.05. Hahnöfer Sand/STD (Rahr 1994). Maxima der Erstbruten werden in der 35. bzw. 37. Pentade erreicht. Es folgen noch zwei Gipfel (20.07.-24.07.) und (09.-13.08.), die sich wohl aus den Pulli der Ersatz- und Zweitbruten ergeben (Abb. 5); die letzte Meldung von einem Nest mit Jungen stammt vom 28.08. Glasmoor/SE (Otto 2008).
Während der Nestlingszeit von 11 – 20 Tagen, je nach Witterung, erfolgt die Beringung im Alter von 7 – 11 Tagen. Hinze beringte die meisten Nestlinge in der 35. Pentade (20.06.-24.06.) sowohl im Berichtsgebiet als auch in Schleswig-Holstein 1955-1961 und 1979-1994; er beringte insgesamt 1.831 Individuen.
Flügge Jungvögel: Die meisten flüggen Jungvögel wurden Ende Juli/Anfang August im Berichtsgebiet beobachtet, s. Abb. 6. Die erste Familie mit flüggen Jungvögeln wurde am 07.06. in den Kirchwerder Wiesen (Heer 2016) gesehen; einen außergewöhnlich späten diesjährigen Jungvogel meldete Stein am 07.10.1971 aus Neu-Wulmstorf. Nach Abzug der Verluste nach dem Ausfliegen überleben nach Jakober & Stauber (1980) ca. 2,7 Jungvögel/Paar.
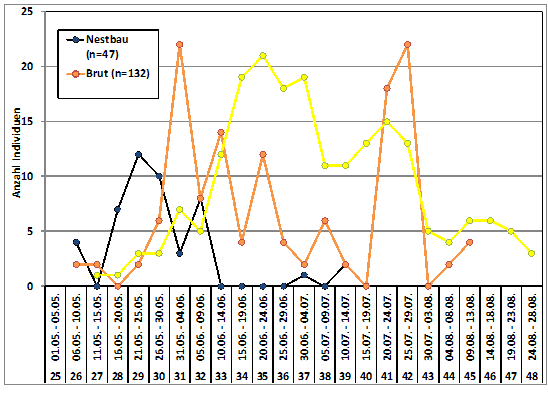
Abb. 5: Neuntöter – Brutaktivitäten im Raum Hamburg
Wegzug
Der Wegzug des Neuntöters aus dem Hamburger Raum beginnt Anfang August und dauert bis Ende September. Gewichtszunahme und Zugunruhe beginnen in Rossitten nach Dolnik (1975) um den 15. August. Gwinner & Biebach (1977) zogen südfinnische und südfranzösische Neuntöter in einem 12-Stunden-Kunsttag auf; bei beiden begann die Zugunruhe Anfang August. Die südfinnischen waren jedoch länger fett. Die früheste Zugmeldung eines vorjährigen Neuntöters stammt vom 12.08. aus dem Süderelbemoor (Nehlsen 1979), die letzte vom 20.10. (Wollin 1963). Ausnahmedaten wie der 20.10.1963 werden von Baum (1969) und Münster (1958) so gedeutet, dass sich der Abzug der Jungvögel bei verspäteten Zweitbruten oder Ersatzgelegen bis in den Oktober verzögern kann.
Letztbeobachtungen:
Zeitraum Median/Spanne
1966-1976; 1987-1995 (n=20): 04.09./13.08.-07.10.
1996-2016: (n=21): 24.09./05.09.-14.10.
1966-1976; 1987-2016 (n=41): 21.09./14.08.-14.10.
Der Median der Letztbeobachtungen hat sich langfristig seit den 1960er Jahren um 21,6 Tage nach hinten verschoben, s. Abb. 7. Zusammen mit den Verfrühungen hat sich damit der Beobachtungszeitraum für die Art seit 1966 um 30,9 Tage vergrößert und beträgt nun im Mittel 135 Tage. In Berlin ist der mittlere Beobachtungszeitraum sogar 150 Tage lang, hat sich aber im Vergleichszeitraum nur um 5 Tage vergrößert, s. Abb. 8. Die Aufenthaltsdauer eines Individuums im Brutgebiet beträgt in Süddeutschland ca. 80 – 100 Tage (Jakober & Stauber 1983).
Verspätung (Median-Letztbeobachtungen)
+ 0,49*** Tage/Jahr (R²= 0,28; p< 0,001) = 21,6 Tage.
Beobachtungszeitraum:
+ 0,71*** Tage/Jahr (R²= 0,40; p <0,001) = 30,9 Tage.
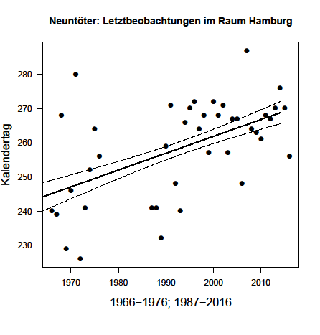
Abb. 7: Neuntöter – Letztbeobachtungen im Raum Hamburg
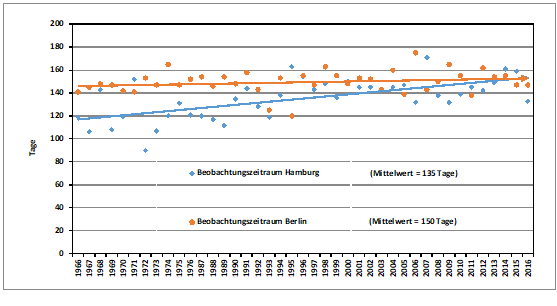
Abb. 8: Neuntöter – Veränderung der Beobachtungszeiträume in Berlin und Hamburg (1966-1976; 1987-2016)
Literatur:
Bairlein, F. & J. Dierschke, V. Dierschke, V. Salewski, O. Geiter, K. Hüppop, U. Köppen, W. Fiedler (2014): Atlas des Vogelzugs. Aula-Verlag Wiebelsheim. 567 S.
Baum, L. (1969): Der Neuntöter (Lanius collurio L.) im Hamburger Raum. – Hamburger avifaun. Beitr. 7: 115-127.
Beiche, S. % I. Luge (2006): Habitatsuswahl und Reproduktion einer Neuntöterpopulation im Nordteil des Köthener Gebietes. Apus 13: 102-123.
Brambilla, M., D. Rubolini & F. Guidali (2007): Between land abandonment and agricultural intensification; habitat preferences of Red-backed Shrike Lanius collurio in low-intensity farming conditions. Bird Study 54: 160-167.
Brandl, R. ,W. Lübcke & W. Mann (1986): Habitatwahl beim Neuntöter Lanius collurio. J. Orn. 127: 69-78.
Bruns, H. (1961): Erstankunft und Sangesbeginn der Vögel in Hamburg 1948-1957. Orn. Mitt. 4: 1-16.
Dietrich, F.. (1928): Hamburgs Vogelwelt. Otto Meißners Verlag, Hamburg, 398 S.
Dolnik, V. (1975): The effects of photoperiods on the timing oft he autumnal migratory start in the Red-backed Shrike (Lanius collurio), an equatorial and transequatorial migrant. Commun. Baltic Commiss. Study Bird Migr. 8: 99-106.
Friedel, W. & S. Ernst (2017 im Druck): 50 Jahre Beobachtung, Dokumentation und Auswertung der Erstankunftstermine heimkehrender Zugvögel im Vogtland – eine Analyse zwischen 1967 und 2016. Mitt. Sächs. Ornithol.
Glutz von Blotzheim, Urs N. & Kurt M. Bauer (1985): Handbuch der Vögel Mitteleuropas, Band 10, Teil 1. Aula-Verlag, 507 S.
Gwinner, E. & H. Biebach (1977): Endogene Kontrolle der Mauser und der Zugdisposition bei südfinnischen und südfranzösischen Neuntötern (L. c.). Vogelwarte 29: 56-63.
Jakober, H. & W. Stauber (1980): Untersuchungen an einer stabilen Neuntöterpopulation (Lanius collurio). J. Orn. 121: 291-292.
Jakober, H. & W. Stauber (1981): Habitatansprüche des Neuntöters Lanius collurio – Ein Beitrag zum Schutz einer gefährdeten Art. Ökol. Vögel 3: 223-247.
Jakober, H. & W. Stauber (1983): Zur Phänologie einer Population des Neuntöters (L. c.). J. Orn. 124: 29-46.
Jakober, H. & W. Stauber (1987): Zur Populationsdynamik des Neuntöters (Lanius collurio). Beih. Veröff. Naturschutz Landschaftspflege Bad.-Württ. 48: 71-78.
Jakober, H. & W. Stauber (1989): Beeinflussen Bruterfolg und Alter die Ortstreue des Neuntöters (Lanius collurio)? Vogelwarte 35: 32-36.
Knysh, N. P. (1994): Influence oft he disturbance factor and inventigations pressure on the nest ruin of passerine bird (a case study oft he Red Backed Shrike). Berkut 3: 38-39.
Kowalski, H. (1995): Wahl der Neststandorte in einer traditionellen Bruthecke des Neuntöters (Lanius collurio). Charadrius 31: 39-41.
Kuzniak, S. (1991): Breeding ecology oft he red-backed Shrike Lanius collurio in the Wielkopolska region (Western Poland). Acta ornithol. 26: 67-84.
Luder, R. (1986): Abnahme der durchschnittlichen Gelegegröße (1901-1977) beim Neuntöter, Lanius collurio, in der Schweiz. Ornithol. Beob. 83: 1-6.
Mann, W. (1987): Ernährungsbiologie des Neuntöters (Lanius collurio). Charadrius 23: 28-38.
Münster, W. (1958): Der Neuntöter oder Rotrückenwürger, Wittenberg-Lutherstadt.
Poltz, W. (1975): Über den Rückgang des Neuntöters (Lanius collurio). Vogelwelt 96: 1-19.
Rohweder, J. (1875): Die Vögel Schleswig-Holsteins. Thomsen. Husum.
Schönfeld, V. (1986): Verbreitung und Habitatwahl des Neuntöters (Lanius collurio) im Rhein-Lahn-Kreis. Ornithol. Naturschutz Regier.Bez. Kobl. 8: 127-132.
Schreurs, T. (1971): Der Rotrückige Würger (Lanius collurio). Paarungshandlungen, Stimmäusserungen und Gelege. Die Heimat, Zeitschr. f. niederrhein. Heimatpflege 42: 69-81.
Schuler, U. (1982): Brutplätze des Rotrückenwürgers an der Autobahn. Vögel der Heimat 52: 197-198.
Simek, J. (201): Patterns of breedind fidelity in the Red-backed Shrike (Lanius collurio). Ornis Fenn. 78: 61-71.
Stein, H. (1972): Über Ankunft und Legebeginn des neuntöters (Lanius collurio) in den Bezirken Magdeburg und Halle. Apus 2: 266-272.
Stresemann,E. (1944): Der zeitliche Ablauf des Frühjahrzuges beim Neuntöter (Lanius collurio). Orn. Mber.52: 1-8.
Tauchnitz, H. (1968): Zur Brutbiologie und Brutdichte des Rotrückenwürgers in einem Auegebiet bei Halle, Apus 1: 229-233.
Tryjanowski, P., s. Kuzniak & B. Diehl (2004): Does breeding performance of Red-backed Shrike, Lanius collurio, depend on nest site selection? Ornis Fenn. 77: 137-141.
Ullrich, B. (1971): Untersuchungen zur Ethologie und Ökologie des Rotkopfwürgers in Südwestdeutschland im Vergleich zum Raubwürger, Schwarzstirnwürger und Neuntöter. Vogelwarte 26: 1-77.
Vanhinsbergh, D. & A. Evans (2002): Habitat associations oft he Red-backed Shrike (Lanius cullurio) i Carinthia, Austria. J. Orn. 143: 405-415.
Vizslan, T. & B. Pingitzer (2000): Data on breeding biology of Red-backed Shrike (Lanius collurio). Aquila 105: 170-171.
Wollscheid, K.-U. (1995): Nistplatzwahl und Habitatnutzung beim Neuntöter (Lanius collurio) aus Beitr. Nat. Kd. Niedersachs. 48: 157-163.
Zang, H. & H. Heckenroth (1998): Die Vögel Niedersachsens, Bartmeisen bis Würger. Naturschutz Landschaftspfl. Niedersachs. B, H.2.10.
Phänologie-AG (H.-H. Geißler, R. Mulsow; veröffentlicht Januar 2018)