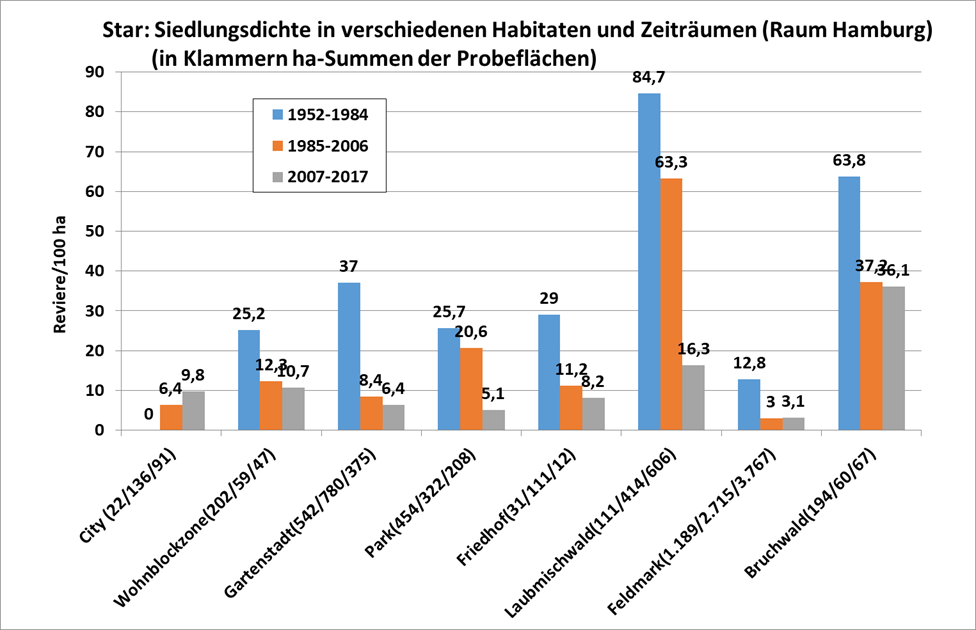
Veränderungen im Jahresrhythmus ausgewählter Zugvogelarten im Raum Hamburg seit den 1960er Jahren
Ronald Mulsow und Luisa Wieczorek
Der Klimawandel und andere Umweltveränderungen beeinflussen auch die Vogelwelt und speziell den Jahresrhythmus von Zugvögeln. In der Literatur findet man am häufigsten Ergebnisse bezüglich Veränderungen von Erstankunftszeiten (Bairlein & Heiser 2014, Bergström & Schütt 2006, Christen 2007, Ernst 2013, Fischer 2002, Hüppop & Hüppop 2002, 2003, 2005, Jenkins & Watson 2000, Kooiker 2005, Moller et al. 2010, Peintinger & Schuster 2005, Rubolini et al. 2007, Schmidt & Hüppop 2007, Schönfeld 2006, Sparks et al. 2007, Tryjanowski et al. 2005, Zalakevicius et al. 2006 u.a.). Seltener sind dagegen Darstellungen zum Einfluss des Klimawandels auf die Zugmuster bei Heim- und Wegzug, auf Brut- und Letztbeobachtungsdaten sowie zu Änderungen beim Beobachtungszeitraum (Hüppop & Hüppop 2005, Schönfeld 2006, Sparks et al. 2007, Witt 2004). Die Auswirkungen des Klimawandels auf die Zyklusphasen während des Aufenthaltes im Brutgebiet werden aber erst deutlich durch die Kombination der Veränderungen in dieser Periode. Ziel dieser Arbeit ist daher, möglichst alle Veränderungen seit den 1960er Jahren im Jahresrhythmus von Zugvögeln im Hamburger Raum zu dokumentieren.
Material und Methode
Ausgewertet wurden die Daten des Arbeitskreises Vogelschutzwarte Hamburg, Daten aus dem Netzportal „ornitho.de“ und der website www.trektellen.nl. Die Beobachtungen wurden von 80-100 Mitarbeitern seit ca. 1950 gesammelt; nach der Einrichtung von „ornitho.de“ waren es ungefähr 120 aktive Ornithologen. Eine für die Auswertung ausreichende Zahl an Beobachtungen war, je nach Art, ab den 1960er Jahren möglich. Ausgewählt wurden Arten, für die möglichst viele Meldungen zu den entsprechenden Zyklusphasen vorlagen; für 25 Arten trifft das zu, davon sind 7 Arten Kurzstreckenzieher (KZ) und 18 Arten Langstreckenzieher (LZ).
Veränderungen bei der Ankunft wurden durch Regressionsanalysen aller Jahre mit ausreichenden Meldungen und dem Vergleich zweier Zeiträume ermittelt. Da es sich bei Erstbeobachtungen oft um sog. „Ausreißer“ handelt, wurde zusätzlich, wenn möglich, die Ankunft des 20. Individuums ausgewertet. Die ermittelten Trends bei der Erstankunft wurden mit Ergebnissen aus anderen Gebieten in Europa verglichen, um etwaige Einflüsse der geographischen Lage zu erkennen.
Entsprechend den Verfrühungen bei der Ankunft waren auch Veränderungen beim Revierverhalten zu erwarten; hier konnten bei den meisten Arten Trend und Umfang der Verfrühungen bei Sanges-/Balzbeginn bzw. bei zwei Arten für den Brutbeginn errechnet werden.
Zwei Zeiträume wurden ebenfalls verglichen, um Verschiebungen von Median und Zuggipfel bei Heim- und Wegzug festzustellen. Dafür wurden entsprechende Pentadensummen der Frühjahrs- bzw. Herbstmeldungen oder Zugbeobachtungen verwendet und/oder systematisch erhobene Daten durch kontinuierliche Beringungen in der Beringungsstation „Die Reit“, wo täglich Vögel vom 30.6 – 6.11. beringt werden. Für Tagzieher boten sich Daten an von Vogelzugplanbeobachtungen; diese werden fast täglich von Ende August bis Anfang November im Hamburger Yachthafen in Wedel/PI in 2-4 Vormittagsstunden erfasst.
Die Letztbeobachtungen lieferte wieder die Datei des Arbeitskreises. Die Veränderungen der Beobachtungszeiträume innerhalb eines Jahres ergaben sich durch die Kombination der Ergebnisse aus den Regressionsanalysen der Erst- und Letztbeobachtungen.
In den späten 1970er und frühen 1980er Jahren gab es eine niedrige Meldehäufigkeit, so dass für einige Arten für diesen Zeitraum keine sinnvolle Auswertung möglich war. Für die meisten Arten ergibt sich aber innerhalb des Zeitraumes 1960-2021 eine Auswertungszeit von 40-60 Jahren (Mittelwert 49,8 Jahre).
Statistische Berechnungen wurden mit dem Rechenprogramm R 4.1.2. (2021)
durchgeführt.
Ergebnisse
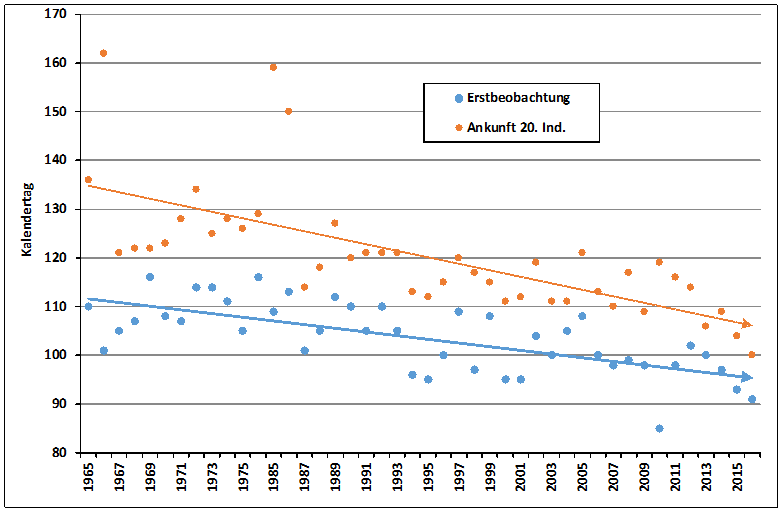
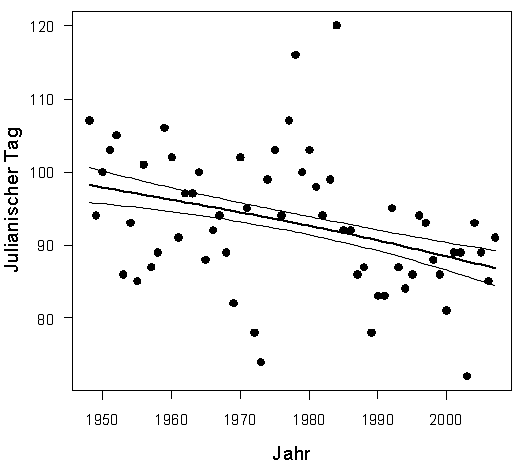
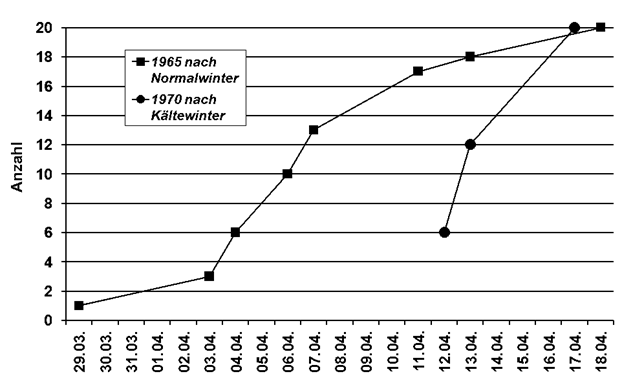
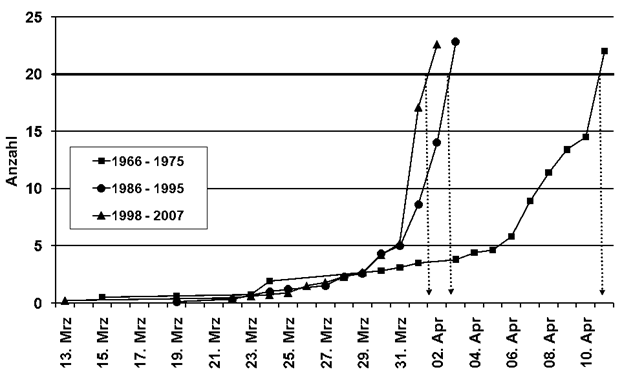
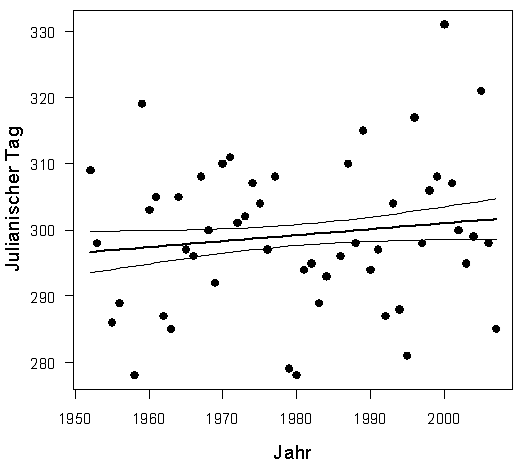
Erstbeobachtung: In Tabelle 1 sind Median-Datum, Spanne und Trend für alle 24 Arten, ausgenommen Kiebitz, (da inzwischen Jahresvogel) angegeben. Außerdem die entsprechenden Ergebnisse aus der Gegenüberstellung von zwei Vergleichs-Zeiträumen. Bei 23 Arten hat sich das Erstbeobachtungsdatum verfrüht; bei 10 Arten signifikant. Lediglich die Gartengrasmücke zeigt eine Verspätung, eine Art, die deutliche Bestandseinbußen in ganz Nordwesteuropa aufweist (Mitschke 2012). Bestandsabnahme und geringe Auffälligkeit könnten bei dieser Art zu einer Verschleierung der wirklichen Erstankunft führen. Nach den Regressionsanalysen betragen die Verfrühungen im Mittel 0,24 Tage pro Jahr (-0,06 bis -1,34), Tab. 1. Am stärksten hat sich die Erstankunft im Gesamtzeitraum bei Hohltaube (-64,2 Tage; in Mecklenburg-Vorp. -62 Tage nach Schmidt & Hüppop 2007), Weißstorch (-43,7 Tage) und Mönchsgrasmücke (-17,3 Tage) verfrüht. Im Vergleich der Zugtypen verfrühte sich die Erstankunft bei Kurzstreckenziehern um 8,6 Tage mehr als bei Langstreckenziehern, im Mittel -18,1/-9,5 Tage. Dieser Unterschied ist schwach signifikant (Mann-Whitney-Test, p =0,03125). Insgesamt findet also eine, in Übereinstimmung mit o.g. Autoren, Verfrühung der Erstankunft statt; wie schon Sokolov et al. (1998) feststellten, ist die Verfrühung besonders ausgeprägt bei den früh heimkehrenden Arten. Nur wenige Autoren stellten keinerlei Verfrühung fest. Hier wurden aber auch wenige Arten (Reichholf 2006, Sjöberg 2006) und/oder nur kurze Zeiträume (Glutz von Blotzheim 2001) ausgewertet.
Die Ankunftsverfrühungen fanden wohl vor allem in der ersten Periode, den 1960er-1990er Jahren, statt; von 24 Arten verfrühten sich 18 Arten, 6 davon signifikant. Im Zeitraum der 1990er-2020er Jahre waren es nur 11 Arten, davon 4 signifikant.
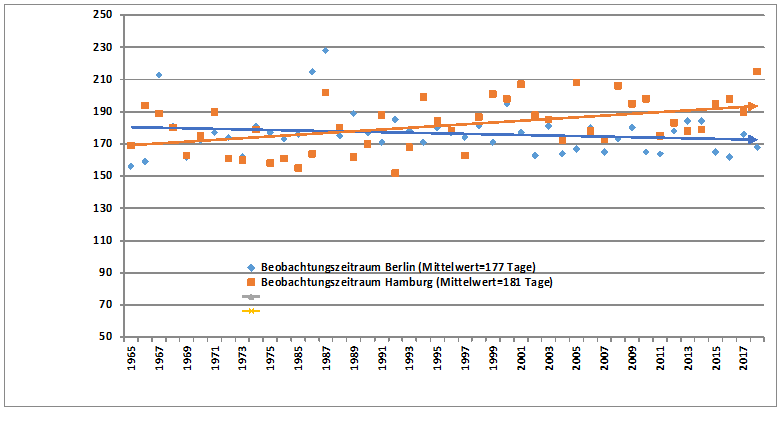
Da die Berliner Ornithologen ihre Erstankunftsdaten jährlich veröffentlichen, war ein zeitidentischer Vergleich mit den Hamburger Daten möglich. Demnach zeigen von 24 Arten 20 eine positive Korrelation, 10 sind signifikant, davon 6 hochsignifikant.
Nach Feststellung einer fast allgemeinen Verfrühung ist die Frage, welche Konsequenzen (Fitness, Reproduktion, Selektion, zeitliche Veränderungen bei Heim- und Wegzug) das für die jeweilige Vogelart mit sich bringt. Wichtig ist demnach die weitere Untersuchung von Bestandsaufbau und Brutgeschehen.
Ankunft des 20. Individuums:
Für 19 Arten waren ausreichende Daten vorhanden, um Trends bei der Heimkehr des 20. Individuums zu berechnen, Tab. 1. Bei allen Arten wurde eine Verfrühung festgestellt, im Mittel 17,9 (5-66) Tage; bei 15 Arten war diese signifikant und meistens auf einem höheren Signifikanzniveau als bei der Erstankunft (Tab. 1). Die 5 Kurzstreckenzieher verfrühten sich im Mittel um 17,3 (8,9-21,7) Tage, alle signifikant; die Langstreckenzieher um 18,1 (5-66) Tage, von 14 sind 10 signifikant. Die Korrelationsberechnung mit den Erstbeobachtungen ergab bei 19 Arten 16 signifikante Werte, davon 13 hochsignifikant. Damit ist belegt, dass nicht nur einige erste Vögel früher ankommen sondern auch größere Anteile der Populationen. Es gilt also auch für Mischpopulationen (Brut- und Zugvögel), was Hüppop & Hüppop (2002, 2005) für Zugvögel auf Helgoland nachweisen konnte. Die Ergebnisse zeigen auch, dass alle Arten früher heimkehren, auch wenn dies bei den Erstankunftsdaten nicht oder kaum deutlich ist.
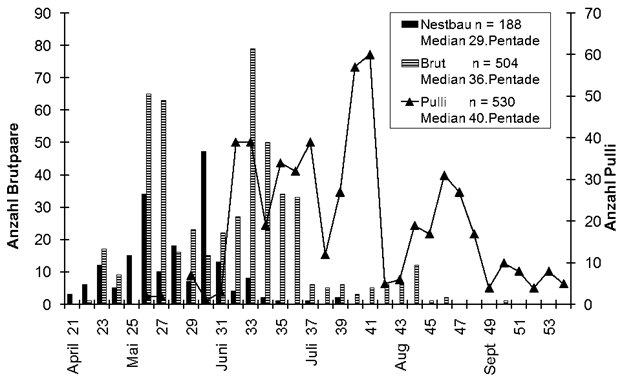
Brutzeitaktivitäten (Tab. 2):
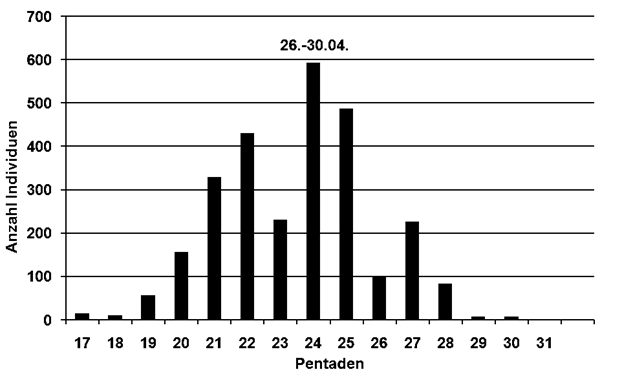
Erwartungsgemäß verfrühten sich auch bei allen 17 untersuchten Arten die ersten Brutzeitaktivitäten, im Mittel um 14,6 Tage (2,0-69,1). Bei zwei Arten gab es genügend Daten, um die Verfrühung des Brutbeginns zu ermitteln. Der Weißstorch beginnt nun 26,1 Tage (48 Jahre innerhalb 1965-2021) früher mit der Brut, der Kiebitz 23,9 Tage (1960-2021).
Bei Balz- bzw. Sangesbeginn zeigten 15 Arten im Mittel eine Verfrühung von 13,2 Tagen (2-69,1). Wie bei der Erstankunft weisen die 5 Kurzstreckenzieher wieder deutlich größere Verfrühungen auf mit durchschnittlich 25,7 Tagen (8,1-69,1). Höchste Werte zeigen Hohltaube (69,1 Tage) und Mönchsgrasmücke (30,5 Tage). Die 10 Langstreckenzieher begannen in den letzten Jahren im Mittel 7 Tage (2-13,3) früher mit dem ersten Revierverhalten; Die stärksten Verfrühungen zeigten Rohrweihe (13,3 Tage), Dorngrasmücke (10,7 Tage) und Baumpieper (10,5 Tage).
Heimzug (Tab. 3):
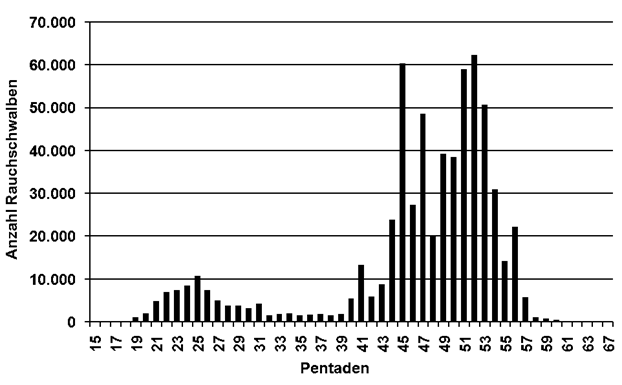
Leider wurde im Frühjahr in Hamburg nicht über einen längeren Zeitraum systematisch beringt und es gibt auch keine Vogelzugplanbeobachtungen. So konnten hier nur Zugmeldungen (4 Arten) und Pentadensummen der Zufallsmeldungen (12 Arten) ausgewertet werden; für den Fitis gab es ältere Beringungsveröffentlichungen (Dinse 1991, Drechsel 1969). Für alle Arten ergab es im Mittel eine Verfrühung des Medians um 1,7 Pentaden (1-4); vier Arten zeigten keine Veränderung. Bei den Kurzstreckenziehern betrug die mittlere Verfrühung 1,6 Pentaden (1-3); bei den Langstreckenziehern 1,8 Pentaden (1-4).
Der Zuggipfel verschob sich bei vier KZ um -4,3 Pentaden (1-7), bei den LZ im Mittel nur um -2,1 Pentaden (1-6). Vier Arten blieben ohne Veränderung. Im Zugmuster des Heimzugs gab es die stärksten Verfrühungen bei Hohltaube, Weißstorch, Wiesenschafstelze und Mönchsgrasmücke.
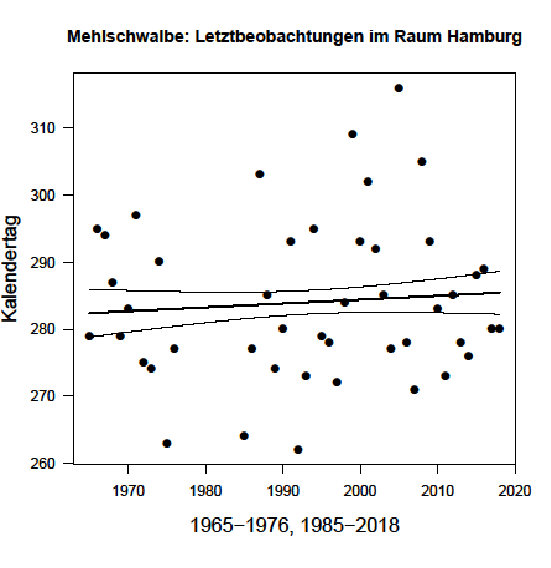
Interessant ist eine Verspätung der Heimzug-Letztbeobachtung beim Fischadler (statt früher Mitte Mai aktuell Ende Mai); die Art ist in Hamburg nur Durchzügler.
Wegzug (Tab. 3):
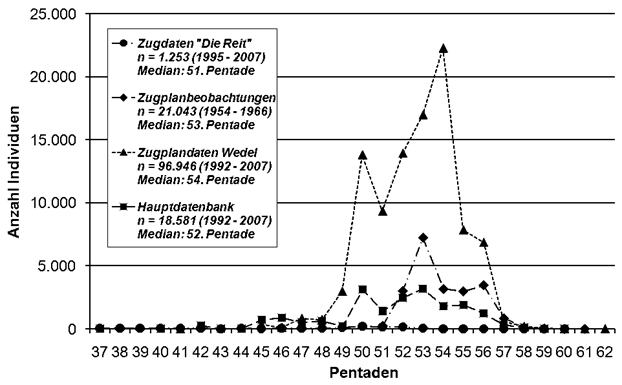
Für 23 Arten konnten folgende Datenquellen genutzt werden: Für 10 Arten standen systematisch erhobene Beringungsdaten zur Verfügung, für 10 weitere gab es ausreichende Zugmeldungen; bei zwei Arten wurden Herbst-Pentadensummen verglichen, für die Bachstelze wurden Daten aus Vogelzugplanbeobachtungen ausgewertet.
Im Gegensatz zu den Heimzugergebnissen sind die des Herbstzuges recht unterschiedlich. Bei den meisten Arten (9) gibt es keine Veränderung des Medians, bei sieben eine Verfrühung und sieben eine Verspätung, im Mittel jeweils um 1,1 Pentaden.
Drei KZ zeigen eine Verspätung um 1 Pentade, die vier anderen keine Veränderung. Bei den LZ verfrüht sich der Median bei sieben Arten, vier LZ verspäten sich, beide im Mittel um 1,3 Pentaden, fünf bleiben ohne Veränderung. Für LZ ist es vorteilhaft, möglichst früh den Sahel zu erreichen, weil dort im Juli-August die Regenzeit für günstige Nahrungsbedingungen sorgt (Lensink 2013).
Bei drei Arten wurden jeweils zwei zusätzliche Vergleichszeiträume ausgewertet. So konnten für die Singdrossel die Beringungsergebnisse durch Daten aus Vogelzugplanbeobachtungen bestätigt werden. Beim Sumpfrohrsänger wurden ältere Beringungswerte durch neuere bestätigt. Unerwartet ergeben die beiden Beringungsauswertungen beim Teichrohrsänger gegensätzliche Ergebnisse; bis 2007 hat sich der Zuggipfel verfrüht, von 2008-2021 dagegen verspätet.
Beim Kiebitz unterscheidet man als Besonderheit den Frühwegzug (Ende Mai bis Ende Juli) vom eigentlichen Herbst-Wegzug (Anfang August bis Mitte Dezember); in den letzten Jahrzehnten hat sich beim Frühwegzug der Zuggipfel um zwei, der Median um eine Pentade verfrüht. Dies war angesichts der Brutverfrühung (Median: 2 Pentaden) zu erwarten, da es sich hier wohl um erste selbständige Jungvögel und gescheiterte Brutvögel handelt. Beim eigentlichen Herbst-Wegzug-Gipfel ergibt sich dagegen, wie bei anderen KZ, eine Verspätung, hier um eine Pentade.
Letztbeobachtungen (Tab. 4)
Seit den 1960er Jahren haben sich die Letztbeobachtungen im Hamburger Raum für 23 Arten verspätet; lediglich der Wespenbussard zeigt eine geringfügige Verfrühung von 2,2 Tagen. Die mittlere Verspätung für alle 24 Arten beträgt 0,363 Tage /Jahr = ca. 18 Tage; sie ist bei 13 Arten signifikant. Die durchschnittlichen Verspätungen sind bei den KZ deutlich größer als bei den LZ:
KZ: 0,528 Tage/Jahr = 25,4 Tage (6-75,7)
LZ: 0,309 Tage/Jahr = 15,5 Tage (-2,2 bis + 38,7).
Höchste Verspätungen zeigen Hohltaube und Zilpzalp, bei den LZ Schwarzmilan und Knäkente.
Beobachtungszeitraum (Tab. 4):
Für alle 24 hier untersuchten Vogelarten ergibt sich durch die Verfrühung der Ankunft und die Verspätungen bei den Letztbeobachtungen eine Verlängerung der Beobachtungszeiträume. Sie beträgt im Mittel 31,3 Tage; 12,9 Tage Verfrühung bei der Ankunft und 18,4 Tage Verspätung beim Abzug.
Auch hier zeigen sich bei den KZ die deutlich höheren Werte:
KZ: 0,907 Tage/Jahr = 43,3 Tage (11,5-139,9)
LZ: 0,554 Tage/Jahr = 27,3 Tage (5,3-70,3).
Die größten Verlängerungen des Beobachtungszeitraumes verbuchen bei den KZ Hohltaube (139,9 Tage) und Zilpzalp (36,4 Tage) für sich, bei den LZ erlangten Schwarzmilan (85,2 Tage) und Weißstorch (70,3 Tage) die höchsten Werte.
Damit ist die Tendenz, zumindest für einzelne Vögel, deutlich; frühere Ankunft im Brutgebiet und späterer Abzug gewähren mehr Zeit für Brutaktivitäten. Über die Zeit der Anwesenheit im Brutgebiet bleibt aber manches im Unklaren, da biologische und methodologische Variabilität zu mehr unsicheren Ergebnissen führen.
Diskussion
Allgemein zeigen die Ergebnisse eine Tendenz zur Reduktion des Zugumfangs bei gleichzeitiger Vergrößerung der Zeiträume; bei den KZ eine Abnahme der ziehenden Vögel und Verkürzung der Zugstrecken, letzteres z.T. wohl auch bei den LZ. Auch Zugrichtungsveränderungen sind möglich, wie das Beispiel Mönchsgrasmücke zeigt (Berthold 1998), ist aber bei anderen Arten bisher kaum nachgewiesen. Vergleicht man die Erstbeobachtungstrends mit Ergebnissen anderer Gebiete in Europa (Moller et al. 2010, Sparks et al. 2005), so liegen die Hamburger Werte z. B. bei den meist untersuchten Arten Fitis und Zilpzalp niedriger. Für die meisten Arten entspricht der „Europawert“ etwa einem Mittel aus Erstbeobachtungstrend und dem Trend der Ankunft des 20. Individuums in Hamburg.
Insgesamt ergibt sich für Heimzug, Ankunft und Brutaktivitäten ein recht einheitliches Phänomen der deutlichen Verfrühungen. Dagegen ist das Herbstbild bezüglich Wegzug und Letztbeobachtungen sehr unterschiedlich und schwieriger zu interpretieren.
Die „Gewinner“ der Gesamtveränderungen scheinen die KZ zu sein; alle Arten mit höheren Ankunftsverfrühungen zeigen im Hamburger Raum deutliche Populationszuwächse (Mitschke 2012). Hausrotschwanz- und Singdrosselbestände sind zumindest stabil. In einem Kartierungsgebiet im Osten Hamburgs hat sich der Bestand des Zilpzalps von 1985-2009 verzehnfacht (R2=0,91), bei der Mönchsgrasmücke stieg er um das Siebenfache (R2=0,86,
Mulsow & Runge 2019). Bestandszunahmen durch eine verlängerte Reproduktionszeit wurden auch in England festgestellt (Newson et al. 2016). Allerdings sind die Beringungszahlen für alle Laubsänger und Grasmücken an der Beringungsstation „Die Reit“ seit ca. 2014 rückläufig (Nahrungsmangel?), womit Zukunftsprognosen wieder ungewiss werden. Andere Umweltveränderungen, z.B. durch die Intensivierung der Landwirtschaft, können aber aus den Gewinnern auch „Verlierer“ machen wie das Beispiel Kiebitz zeigt. In dem o.g. Kartierungsgebiet fiel der Bestand in den 25 Jahren von 24 Revieren auf 0 (R2=0,80).
Die genetisch stärker fixierten LZ sind wegen des geringeren Zeitgewinns benachteiligt gegenüber den KZ, was sich bei vielen Singvogelarten in Bestandsabnahmen zeigt. Bei früh heimkehrenden Arten (Weißstorch, Rohrweihe) zeigen sich meist größere Verfrühungen, bei späten Heimkehrern (Mauersegler, Kuckuck, Klappergrasmücke) dagegen geringere. Hier spielt aber auch Zugwegverkürzung z. T. eine wichtige Rolle, nachgewiesen z.B. beim West-Weißstorch (Schulz 2019), inzwischen vereinzelt auch bei Ost-Weißstörchen (Mitt. Thomsen, Storchendorf Bergenhusen 2022). Rohrweihenweibchen und –jungvögel überwintern aktuell teilweise in SW-Europa (Bairlein et al. 2014), also näher zum Brutgebiet hin (Bairlein et al. 2014).
Da die Regenzeit im Sahel in Juli/August stattfindet, bringt es für LZ Selektionsvorteile, möglichst früh in Europa abzuziehen, um das entsprechende Nahrungsangebot nutzen zu können. Zehn von Fünfzehn Arten zeigen eine Verfrühung des Wegzugmedians bzw. –gipfels um 1-3 Pentaden als Hinweis auf dieses Verhalten. Fünf Arten, darunter auch Baumfalke und Wespenbussard, verspäten sich jedoch um 1-2 Pentaden, ein Verhalten, das ungeklärt bleibt. Rätselhaft bleibt auch eine Verfrühung des Wegzugmusters beim Teichrohrsänger bis 2007, danach aber eine Verspätung, jeweils um eine Pentade. Für diese Art werden aktuell vermehrt Überwinterungsversuche in Nordafrika und Spanien gemeldet (Bairlein et al. 2014), was eine deutliche Zugwegverkürzung bedeutet. Die relativ große Verfrühung bei der Ankunft könnte auch zu einer Vermehrung der Bruten pro Saison führen.
Das unterschiedliche Verhalten der LZ beim Wegzug führt zu der Frage: Was bringt der Art mehr Selektionsvorteile: Mehr Bruten in Europa oder „rechtzeitige“ Ankunft in Afrika im Sahel? Möglicherweise profitierten beim Teichrohrsänger seit 2007 Populationen, die die erstere Option bevorzugten. Wegen des sehr langen Zugweges blieb der Sumpfrohrsänger dagegen bei der alternativen Strategie. Hier gilt es zu klären, ob alle „Ein-Brut – Arten“ eine Verfrühung beim Wegzug wählen (Jenni & Kery 2003). Bei Wespenbussard und Baumfalke könnte wegen der schwachen Datengrundlage ein falsches Ergebnis vorliegen.
Finden größere Umweltveränderungen statt, scheint es sinnvoll, die genetische Reaktionsnorm voll auszunutzen. Möglicherweise ist so die Tatsache zu erklären, dass auch bei den LZ der „letzte Vogel“ immer später abzieht. KZ, die als weniger genetisch fixiert gelten, können da noch länger auf günstigen Rückenwind oder besseres Wetter warten (Haest et al. 2019). Die Arten mit den höchsten Trends bei den Letztbeobachtungen (Hohltaube, Zilpzalp, Hausrotschwanz) fielen in den letzten Jahren durch immer häufigere Überwinterungsversuche auf. Insgesamt bestätigen die Ergebnisse also eine Zunahme der Anteile von Teilziehern und Standvögeln. Weitere langfristig wirkende Einflussfaktoren wie die Nahrungswahl, Abnahme der Insekten und Verhalten im Überwinterungsgebiet müssten aber noch untersucht werden.
Insgesamt scheint die Vogelwelt auf die einschneidenden Umweltveränderungen im Anthropozän mit einer Vergrößerung der phänotypischen Variabilität zu reagieren.
Danksagung:
Für das Bereitstellen der Datengrundlagen bedanken wir uns herzlich bei dem Arbeitskreis Vogelschutzwarte Hamburg. Beim Erstellen von Abfragen und Problemlösen von Informatikfragen war Herr Prof. Dr. E. Fähnders hilfreich. Bei den statistischen Auswertungen wurden wir effektiv beraten und unterstützt von Herrn Dr. O. Hüppop, der uns außerdem wichtige projektspezifische Literatur zukommen ließ.
Für Manuskriptdurchsicht und Korrekturen haben wir Frau M. Mulsow und Herrn J. Berg zu danken. Gefördert wurde das Projekt durch die Deutsche Ornithologische Gesellschaft.
Literatur:
Bairlein F, Dierschke J, Dierschke V, Salewski V, Geiter O, Hüppop K, Köppen U, & Fiedler W 2014: Atlas des Vogelzugs. Aula-Verlag Wiebelsheim.
Bairlein F & Heiser F 2014: Langfristige Veränderungen in der Frühjahrsankunft von Zugvögeln im Lech-Donau-Winkel, Bayern. Ornithol. Anzeiger 53: 1-21.
Bergström E. & Schütt L 2006: En langtidstudie (1938-204) av flyttfaglarnas ankomst til mellersta Värmland. Ornis svecica 16: 95-111.
Berthold P 1998: Vogelwelt und Klima: Gegenwärtige Veränderungen. Naturw. RDsch. 51: 337-348.
Christen W 2007: Erstankunft ausgewählter Zugvogelarten bei Solothurn. Ornithol. Beobachter: 104:53-63
Dinse V 1991: Über den Heimzug von Kleinvögeln in Hamburg. Eine Auswertung im Rahmen des Mettnau-Reit-Illmitz-Programms. Hamburger avifaun. Beitr. 23: 1-125.
Drechsel H. 1969: Die Hohe Reit (Hamburg-Vierlande) als Beringungsgebiet. Hamburger avifaun. Beitr.7: 1 – 11.
Ernst S. 2013: Veränderungen der Ankunftszeiten von 25 häufigen Zugvogelarten im sächsischen Vogtland in den Jahren 1967 bis 2011. Mitt. Ver. Sächs. Ornithol. 11: 1-14.
Fischer S 2002: Frühjahrsankunft ziehender Singvogelarten in Berlin über 26 Jahre. Berl. ornithol. Ber. 12: 145-166.
Glutz von Blotzheim U N 2001: Phänologie der häufigsten Brutvögel auf Sturmwurf- und Jungwuchsflächen im Tannen-Buchenwaldareal am Schwyzer Nordalpenrand (750-1200 m ü. M.). Ornithol. Beob. 98: 113-134.
Haest B, Hüppop O, van de Pol M, & Bairlein F 2019: Autumn bird migration phenology: A potpourri of wind, precipitation and temperature effects. Glob Change Biol. 2019; 00: 1-17.
Hüppop O & Hüppop K 2002: North Atlantic Oscillation and timing of spring migration in birds. Proc. R. Soc. Lond. B (2003) 270: 233-240.
Hüppop O & Hüppop K 2003: North Atlantic Oscillation and timing of spring migration in birds. Proc. R. Soc. Lond. B 270: 233-240.
Hüppop K & Hüppop O 2005: Atlas zur Vogelberingung auf Helgoland. Teil 3: Veränderungen von Heim- und Wegzugzeiten von 1960 bis 2001. Vogelwarte 43: 217-248.
Jenkins D & Watson A 2000: Dates of first arrival and song of birds during 19784.99 in mid-Deeside, Scotland. Bird Study 47: 249-251.
Jenni L & Kery M 2003: Timing of autumn bird migration under climate change: advances in long-distance migrants, delays in short-distance migrants. Proc. R. Soc. Lond. DOI 10.1098.2003.2394
Kooiker G 2005: Vögel und Klimaerwärmung: 28-jährige phänologische Beobachtungen in und um Osnabrück von 1976 bis 2004. Vogelkundl. Ber. Niedersachs. 37: 99-111.
Lensink R, Troost G & Pilzecker J 2013: Aankomst, doortrek en vertrek van de Huiszwaluw, Delichon urbicum in Nederland in relatie tot een opwarmend klimaat. Het Vogeljaar 61: 155-164.
Mitschke A 2012: Atlas der Brutvögel in Hamburg und Umgebung. Hamburger avifaun. Beitr. 39: 5-228.
Moller A P, Fiedler W & Berthold P 2010: Effects of climate change on birds. Oxford, University Press.
Mulsow R & Runge U 2019: Zur Bestandsentwicklung der Brutvogelwelt im Naturschutzgebiet Stellmoorer Tunneltal. Hamburger avifaun. Beiträge 44: 16-57.
Newson S E, Moran N J, Musgrove, A J, Pearce-Higgins, P, Gillings, S, Atkinson P, Miller, W, Grantham R & Baillie S R 2016): Long-term changes in the migration phenology of UK breeding birds detected by largescale citizen science recording schemes. Ibis 158: 481-495.
Peintinger M. & S Schuster 2005: Veränderungen der Erstankünfte bei häufigen Zugvogelarten in Südwestdeutschland. Vogelwarte 43: 161-169.
Reichholf H. 2006: Zilpzalp Phylloscopus collybita und Fitis Ph. trochilus in der Isaraue bei München: Erstankunft, Frühjahrszug und Brutzeit-Präsenz. Orn. Mitt. 58: 45-52.
Rubolini D, Moller A P, Rainio K & Lehikoinen E 2007: Intraspecific consistency and geographic variability in temporal trends of spring migration phenology among European bird species. Climate Research 35: 135-146.
Schmidt E & Hüppop K 2007: Erstbeobachtung und Sangesbeginn von 97 Vogelarten in den Jahren 1963 bis 2006 in einer Gemeinde im Landkreis Parchim (Mecklenburg-Vorpommern). Vogelwarte 45: 27-58.
Schönfeld M 2006: Medianwerte der Erst- und Letztfeststellungen ausgewählter Zugvögel im Vergleich von 1975 bis 2005 sowie von Zehnjahreszeiträumen für den Altkreis Wittenberg/Sachsen-Anhalt. Orn. Mitt. 58: 131-140.
Schulz H. 2019: Boten des Wandels. Rowohlt Polaris
Sjöberg N 2006: Ankomstdatum 1948-2006 och häckning 1978-2006 i Jämtland för svartvit flugsnappere Ficedula hypoleuca, rödstjärt Phoenicurus
phoenicurus och lövsangere Phylloscopus trochilus. Ornis svecica 16: 118-126.
Sokolov L V, Marcovets M Y, Shapoval A P & Y. Morozov G 1998: Long-term trends in the timing of spring migration passerines on the Courish Spit of the Baltic Sea. Avian Ecology and behaviour 1: 1-21.
Sparks T H, Bairlein F, Bojarinova J G, Hüppop O, Lehikoinen E A, Rainio, K, Sokolov L V & Walker D 2005:11, 22-30: Examining the total arrival distribution of migratory birds. doi: 10.1111/j.1365-2486.2004.00887.x.
Sparks T H, Huber K, Bland R L, Crick H Q P, Croxton P J, Flood J, Loxton R G, Mason C F, Newnham J A & Tryjanowski P 2007: How consistent are trends in arrival (and departure) dates of migrant birds in the UK? J. ornithol. 148: 503-511.
Tryjanowski P., Kuzniak S & Sparks T H 2005: What effects the magnitude of change in first arrival dates of migrant birds? J. Ornithol. 146: 200-205.
Zalakevicius M., Bartkeviviene G, Raudonikis L & Janulaitis J 2006: Spring arrival response to climate change in birds: a case study from eastern Europe. J. Orn. 147:326-343.
Witt K. 2004: Erst- und Letztbeobachtungen des Mauerseglers (Apus apus) in Berlin. Berl. ornithol. Ber. 14: 186-192.
Datenquellen:
Beobachtungsdatei des Arbeitskreises Vogelschutzwarte Hamburg (incl. Daten aus „ornitho.de).
Website: www.trektellen.nl
Zusammenfassung:
Nach zahlreichen Veröffentlichungen über Veränderungen der Erstankunft von Zugvögeln versuchten wir anhand der Beobachtungsmeldungen im Raum Hamburg zu klären, welche Veränderungen im gesamten Jahresrhythmus auftreten. Für diese Arbeit wurden die Daten von 14 Singvogel- und 11 Nichtsingvogelarten aus dem Zeitraum ca. 1960-2021 ausgewertet. Sie beziehen sich auf das Berichtsgebiet des Arbeitskreises Vogelschutzwarte Hamburg (Hamburg und Umgebung = 2.122 qkm). Die Artenauswahl richtete sich nach der Zahl der Meldungen, die für die vorgegebene Fragestellung zur Verfügung standen. Veränderungen im Jahresrhythmus wurden durch Regressionsanalysen und Vergleiche zweier Zeiträume ermittelt.
Ergebnisse: Eine Verfrühung der Erstbeobachtungen zeigt sich bei 23 von 24 Arten. Die einzige Verspätung bei der Gartengrasmücke wird eventuell durch starke Bestandsabnahmen vorgetäuscht. Größte Verfrühungen gab es bei Hohltaube, Weißstorch und Mönchsgrasmücke. Im Mittel betrug die Verfrühung für alle Arten 0,26 Tage/Jahr, für Kurzstreckenzieher (KZ) 18,1 Tage, für Langstreckenzieher (LZ) 10,2 Tage; der Unterschied ist schwach signifikant. Nach den Vergleichen zweier Zeiträume fanden bei den ausgewählten Arten die Verfrühungen vorwiegend im älteren Zeitraum, 1960er – 1990er Jahrs, statt. Der Vergleich mit Zeitserien aus Berlin ergab überwiegend positive Korrelationen.
Eine deutliche Verfrühung bei der Ankunft des 20. Individuums wurde bei allen Arten festgestellt. Erstes revieranzeigendes Verhalten wie Erstgesang oder Balz verfrühte sich erwartungsgemäß bei allen untersuchten Arten im Mittel um 13,2 Tage; deutlicher wieder bei den KZ. Bei Weißstorch und Kiebitz war eine deutliche Verfrühung des Brutbeginns nachweisbar.
Beim Heimzugmuster zeigte sich, trotz relativ schwacher Datenlage, eine allgemeine Verfrühung des Medians bei allen Arten um 1,7 Pentaden, außer bei der Knäkente.
Im Gegensatz zum Heimzug sind die Herbstzug-Auswertungsergebnisse recht unterschiedlich. Neun Arten zeigen keine Veränderung, sieben eine Verfrühung und sieben eine Verspätung des Medians. Beim Teichrohrsänger
ergeben die Beringungszahlen sogar eine Trendumkehr; bis 2007 eine Verfrühung, danach eine Verspätung. Möglicherweise ist es für Arten mit nur einer Jahresbrut vorteilhafter frühzeitig im Sahel anzukommen, um die Vorteile
der Regenzeit zu nutzen, während andere dazu tendieren, ihre Fitness durch mehr Bruten zu erhöhen.
Letztbeobachtungen: Die mittlere Verspätung bei 23 von 24 Arten beträgt 18 Tage; sie ist bei KZ wieder größer. Nur der Wespenbussard verfrüht sich um 2,2 Tage.
Beobachtungszeitraum: Die mittlere Verfrühung der Erstbeobachtungen (12,9) und die mittlere Verspätung der Letztbeobachtungen ergeben eine Verlängerung des Beobachtungszeitraumes um 31,3 Tage.
Die Möglichkeit zu mehr Bruten wird von KZ genutzt und führte zumindest über längere Zeiträume zu Bestandszunahmen. Bei den LZ beeinflussen vermutlich außer dem Klimawandel artspezifisch wirksame Habitatveränderungen im Brut- oder auch Überwinterungsgebiet die Selektion in die eine oder andere Richtung.